实验五 细胞凝集反应(2学时 验证性实验)
一、实验目的
1、观察血细胞的凝集现象;
2、掌握凝集素促使细胞凝集的原理。
二、实验原理
细胞质膜是由蛋白质不同程度镶嵌在脂双层中所形成的动态流动结构,蛋白质和脂类分子又与寡糖链结合为糖蛋白和糖脂分子,糖蛋白和糖脂分子伸至细胞表面的分枝状寡糖链在质膜表面形成细胞外被(又称为糖萼)。许多研究结果表明:细胞间的分子识别、细胞的生长和分化、免疫反应和肿瘤发生等均与细胞外被(分枝状寡糖链)有关。凝集素能与细胞外被的糖分子相连接、在细胞间形成“桥”,从而达到细胞凝集的效果。
凝集素(1ectin)是一类含糖的(少数凝集素例外)并能与糖进行专一性结合的蛋白质,它具有凝集细胞和刺激细胞分裂的作用。凝集素促使细胞凝集主要是由于它能与细胞外被的糖分子相连接、在细胞间形成“桥”的结果,且加入与凝集素互补的糖分子可以抑制细胞间的凝集反应。
三、实验材料及用品
? 实验材料:
1.土豆块茎。
2.2%的红细胞:以无菌方法抽取兔子静脉血液(加抗凝剂),用生理盐水洗5次,每次2000 r/min,离心5 min,最后按沉淀压积的红细胞体积用生理盐水配成2%红细胞悬液
? 器材:显微镜、托盘天平、载玻片若干、滴管2支、离心管2支。 ? 试剂:PBS缓冲液:称取NaCl 7.2g、NazHP04 1.48g、KH2P04 0.43g、加蒸馏水,定容至l000 mL,调pH值到7.2。
四、实验步骤
①称取土豆去皮块茎2g
②加10 mL PBS缓冲液,浸泡2h(浸出的粗提液中含有可溶性土豆凝集素) ③载玻片上滴一滴土豆凝集素
④再滴一滴2%红细胞液,充分混匀
⑤静置20 min
⑥低倍显微镜下观察血球凝集现象
⑦以PBS液加2%血细胞悬液作为对照实验
五、实验作业
1.绘制简图,表示血细胞的凝集过程;
2.图示血细胞凝集的原理。
实验六 细胞膜的通透性观察(2学时 验证性实验)
一、实验目的
1.了解细胞膜对物质通透性的一般规律。
2.了解溶血现象及其发生机制。
二、实验原理
细胞膜是细胞与环境进行物质交换的选择通透性屏障。它是一种半透膜,可选择性控制物质进出细胞。各种物质出入细胞的方式是不同的,水是生物界最普遍的溶剂,水分子可以按照物质浓度梯度从渗透压低的一侧通过细胞膜向渗透压高的一侧扩散,这种现象就是渗透。渗透作用是细胞膜的主要功能之一。将红细胞放在低渗盐溶液中,水分子大量渗到细胞内,可使细胞胀破,血红蛋白释放到介质中,由不透明的红细胞悬液变为红色透明的血红蛋白溶液,这种现象称为溶血。将红细胞放在某些等渗盐溶液中,由于红细胞膜对各种溶质的通透性不同,膜两侧的渗透压平衡会发生改变,也会发生溶血现象。因此,发生溶血现象所需时间长短可作为测量物质进入红细胞速度的一种指标。本实验选用红细胞作为细胞膜透性的实验材料,将其放入不同的介质溶液中,观察红细胞的变化。
三、实验材料及用品
? 材料:鸡血细胞
? 器材:普通显微镜、普通离心机、扭力天平、10ml 试管、10ml 刻度离心管、试管架,2ml 注射器(无需针头)或5ml 移液管、洗耳球、滴管、载玻片、擦镜纸、记号笔等。
? 试剂:
1.Alsever 溶液
葡萄糖 2.05g 柠檬酸钠(Na9C6H5O7.2H2O) 0.89g 柠檬酸(C6H 6O7.H2O) 0.05g 氯化钠 0.42g 蒸馏水 100ml 调PH 至7.2,过滤灭菌或高压灭菌10min,置4℃冰箱保存。
2. 0.128 mol/L NaCl 溶液。
3. 0.05 mol/L NaCl 溶液。
4. 0.4 mol/L NaCl 溶液。
3. 0.128 mol/L NH4Cl 溶液。
4. 0.128 mol/L NH4AC 溶液。
5. 0.128 mol/L NaNO3溶液。
7. 0.32 mol/L 葡萄糖。
8. 0.32 mol/L 甘油。
9. 0.32 mol/L 乙醇。
10. 0.32 mol/L 丙酮。
11. 蒸馏水。
四、实验步骤:
1.从集市买一只活鸡现杀取血5ml(防止污染),放入盛有20ml Alsever’s 液瓶中,混匀后置冰箱保存备用(2 周内使用)。
2. 使用前,取用Alsever’s 液保存的新鲜鸡血5ml , 加入8ml 的0.128 mol/L 的NaCl 溶液,小心混匀,1000 r/min 离心5min,如此3 次洗涤,最后配成30%的鸡红细胞(CRBC)悬液。
3. 取11 支试管,按附表中所示测试溶液,分别取样各3ml,作出标记后,各管均加入红细胞悬液2 滴,混匀后静置于温室中,观察各支试管中发生溶血的时间及其变化。
五、观察结果
1. 试管内液体分两层:上层浅黄色透明,下层红色不透明为不溶血(-),镜检红细胞完好呈双凹盘状。
2. 如果试管内液体混浊,上层带红色者,称不完全溶血(+或++),镜检有部分红细胞呈碎片。
3. 如果试管内液体变红而透明者,称完全溶血(+++),镜检发现细胞全部呈碎片。
六、实验作业
表各种溶液的溶血现象观察结果
编号 测试溶液 是否溶血 时间 分析原因 1 0.128 mol/L NaCl
2 0.05 mol/L NaCl
3 0.4 mol/L NaCl
4 0.128 mol/L NaNO3
5 0.128 mol/L NH4Cl
6 0.128 mol/L NH4AC
7 0.32 mol/L 甘油
8 0.32 mol/L 乙醇
9 0.32 mol/L 丙酮
10 0.32 mol/L 葡萄糖
11 H2O
书写实验报告,并就细胞膜对不同物质的渗透性不同,对实验结果进行分析见表。
目测法判断标准:快速溶血:+++;慢速溶血:++或+;不溶血:-
第二篇:细胞遗传学
第一章 绪 论
第一节 细胞遗传学的研究对象与任务
细胞遗传学=细胞学+遗传学
细胞遗传学是遗传学与细胞学相结合的一个遗传学分支学科。研究对象主要是真核生物,特别是包括人类在内的高等动植物。着重研究分离、重组、连锁、交换等遗传现象的染色体基础,以及染色体畸变和倍性变化等染色体行为的遗传学效应。
细胞遗传学的主要研究对象 染 色 体 能用碱性染料染色、有结构的线状体, 是遗传物质基因的载体。在生物的细胞核中,有一种易被碱性染料染上颜色的物质,叫做染色质。染色体只是染色质的另外一种形态。它们的组成 成分是一样的,但是由于构型不一样,所以还是有一定的差别。
细胞遗传学的任务:
研究染色体的数目、形态、结构、功能与运动,以及这些特征的各类变异对遗传传递、重组、表达与调控的作用与影响。
各自在动植物生殖细胞的减数分裂过程中发现了染色体行为与遗传因子行为之间的平行关系,认为孟德尔所设想的遗传因子就在染色体上,这就是所谓的萨顿-博韦里假说或称遗传的染色体理论。
细胞遗传学衍生的分支学科:
体细胞遗传学--主要研究体细胞,特别是离体培养的高等生物体细胞的遗传规律;
分子细胞遗传学--主要研究染色体的亚显微结构和基因活动的关系;
进化细胞遗传学--主要研究染色体结构和倍性改变与物种形成之间的关系;
细胞器遗传学--主要研究细胞器如叶绿体、线粒体等的遗传结构;
医学细胞遗传学--这是细胞遗传学的基础理论与临床医学紧密结合的新兴边缘科学,研究染色体畸变与遗传病的关系等,对于遗传咨询和产前诊断具有重要意义。
< Cytogenetics and Genome Research> <Genome><Chromosome Research> <Chromosoma> <Genetics> < Genome Research> < The Plant Cell> <Theory and Applied Genetics> <遗传> 《遗传学报》
第二章 染色体的形态结构 一、基本概念
染色质(Chromatin):在尚未分裂的细胞核中,显微镜下可见的可被碱性染料染色较深的、纤细的网状物。
染色体(Chromosome):染色体是遗传物质(DNA)与蛋白质按一定方式结合成核小体,由核小体相连成丝状染色质(Chromatin)再经多重螺旋化形成的具有特定形态结构的一种细胞器。
染色体是核中重要和稳定的成分,具有特定形态结构,具有自我复制能力,积极参与代谢活动,出现连续而有规律性的变化。
染色体控制生物的遗传性状,它在形态、数目、结构上的变化,也会导
致遗传性状的相应变化。 二、研究染色体形态最适合的时期
1. 2.
1.1 有丝分裂中期可以观察到哪些形态特征?
? 次缢痕
? 臂比
? 着丝粒
? 长度
? 数目
1.2 有丝分裂中期染色体的一般形态
(1) 染色体长度:差别悬殊。 0.5μm-30μm
小麦:染色体平均长度11.2 μm, 总长235.4μm。
绝对长度:显微镜下染色体直接测量的长度。一般为1~25μm。最短的长度为0.25μm,最长的长度是30μm。
相对长度:某一染色体绝对长度占该染色体组绝对长度的百分数。 在细胞周期中,染色体处于动态的收缩过程中。
(2)染色体数目
正常情况每种生物不同时期的染色体数目是恒定的。不同物种间染色体数目差异很大。
马蛔虫2条;真蕨纲瓶尔小草属的高达800~1200条。
染色体大、数目少的物种是经典细胞遗传学研究的优良实验材料。 如果蝇、玉米、蚕豆、洋葱、麦类。
3)? 染色体分离的一种“装置”。也是姐妹染色单体在分开前相互联结的位置,在染色体的形态上表现为一个缢痕(constriction)。
? 着丝粒位于异染色质区内,这里富集了卫星DNA,也就是短的DNA串联重复序列。
? 在缢痕区内有一个直径或长度为400 nm左右的很致密的颗粒状结构,这称为动粒(kinetochore)的结构直接与牵动染色体向两极移动的纤丝蛋白相连结。
着丝点(Kinetochore)-着丝粒的外层结构,是细胞分裂时纺锤体微管附着部位.
按着丝粒位置将染色体分为几种类型
1)中着丝粒染色体2)近中着丝粒染色体3)亚中着丝粒染色体4)亚端着丝粒染色体5)近端着丝粒染色体6)端着丝粒染色体
臂比(arm ratio,A)=长臂/短臂(q/p或L/S)
着丝粒指数(Centromeric Index,C)=短臂长度(p)/染色体长度(p+q)×100%
(4)核仁组织区(Nucleolus Organizer Region, NOR)
次级缢痕(副缢痕;核仁形成区) (secondary constriction): 主缢痕外着色较浅的染色体缢缩区,不能弯曲,与核仁形成有关。常在短臂出现。位置相对稳定。 NOR是核糖体RNA合成的场所,植物中18S-5.8S-28SRNA 在该区域转录,5SrRNA在染色体组其他区域转录。 每个物种至少有一对同源染色体具有NOR, 玉米的NOR为一异固缩的疖。
核仁竞争:杂种特别是远缘杂种中,一物种的NOR活 性受到抑制、不能表达的现象。
三级缢痕(Tertiary constriction)
5)随体 (Satellite, SAT)
从次缢痕到臂末端有一种圆形或略呈长形的染色体节段。
1.1减数分裂前期I粗线期可观察到哪些形态特征?
? 疖
? 端粒
? 染色粒
? 常染色质 和异染色质
? 次缢痕
? 臂比
? 着丝粒
? 长度
? 数目
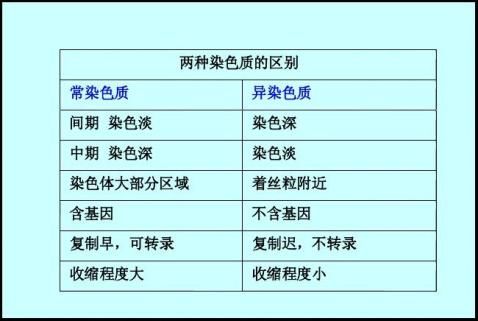
1.2 常染色质(Euchromatin)和异染色质(Heterochromatin)
异固缩:在细胞周期中,某些染色体的某些部分在固缩程度和染色性质上与其他染色体或染色体其他部分不同步的现象。显微镜下观察染色质着色不均匀、深浅不同.
正异固缩—在间期、前期固缩化过程较其它染色质早、染色深 负异固缩—在中、后期固缩化过程较其它染色质迟、染色浅 常染色质(euchromatin):
固缩化过程与细胞周期一致。用碱性染料染色较异染色质区染色浅,一般由单拷贝或寡拷贝DNA组成,具有转录活性;在分裂间期,呈
高度分散状态,DNA合成在S期的早、中期。
异染色质(heterochromatin):
具有异固缩特性的染色质。
含两类:
组成型或结构型异染色质(constitutive heterochromatin)
在任何情况下均表现异固缩特征。一般位于着丝粒、核仁组织形成区和端粒附近。也可出现在其它区域。
兼性或功能型异染色质(facultative heterochromatin)
具有常染色质的组成,在特定组织或特定时期表现异染色质特性。即异染色质化的常染色质,很可能与基因表达、调控有关。
第三章 染色体的物质组成及组装
第一节 原核生物染色体
1、类核体
2、环状染色体:电子显微镜观察
3、物质组成
DNA:环状DNA, 质粒DNA
RNA: RNA病毒,初生转录体
蛋白质:Hu蛋白
2. 原核生物的染色体结构
类核小体模型:
4个蛋白分子
DNA
第二节 真核染色质的结构
染色质(Chromatin)具有压缩结构,其中的大部分DNA 序列结构上难以接近且无功能活性,仅少数是活性序列。染色质的一般结构如何?活性序列与非活性序列之间又有什么区别?有关基因物质高包装比(Packing ratio)的研究表明,DNA 不能被直接包装成染色质的最终结构,因而必然存在分级包装过程。
真核染色体较原核染色体:被核膜包围,形态大,至少2条。
一、真核染色质的组成成分
由DNA、组蛋白、非组蛋白及少量RNA组成,比例为1:1:(1-1.5):0.05。DNA与组蛋白的含量比较恒定,非组蛋白的含量变化较大,RNA含量最少。
1、DNA: 双链DNA, 双螺旋结构 染色体的一级结构
2、 RNA: rRNA
3、蛋白质:
组蛋白(HP, histone protein):
核心组蛋白(core histone)高度保守,包括:
H2a: 赖氨酸含量略高
H2b: 赖氨酸含量略高
H3: 富精氨酸
H4: 富精氨酸
H1: 连接组蛋白(linker histone),富赖氨酸, 具有属特异性和 组织特异性,所以H1是多样性的。
非组蛋白(NHCPs, non-histone chromosomal protein): DNA特异结合蛋白, 包括: 结构蛋白、酶蛋白、调控蛋白、激素受体、高迁移族(HMG)蛋白等。包含染色质中除组蛋白外的所有蛋白质。
非组蛋白的特性:①含有较多的天门冬氨酸、谷氨酸,带负电荷,属酸性蛋白质;②整个细胞周期都进行合成,不象组蛋白只在S期合成,并与DNA复制同步进行;③能识别特异的DNA序列,识别信息存在于DNA本身,位点在大沟部分,识别与结合籍氢键和离子键。
非组蛋白的功能:①帮助DNA分子折叠,以形成不同的结构域,从而有利于DNA的复制和基因的转录;②协助启动DNA复制;③控制基因转录,调节基因表达。
4、无机离子: K+ Cl- Na+ Mg2+ Ca2+
二、核小体(Nucleosome)模型
Olins 夫妇和Chambon在电子显微镜下观察到间期核直径约11nm颗粒,它们与直径1.5-2nm的纤维相连接形成串珠结构, 11nm颗粒称之为核小体。
用非特异性核酸酶(如微球菌核酸酶)处理染色质,大多数情况下可得到大约200bp的片段,但处理裸露的DNA分子会得到随机降解的片段。以这个实验为基础,Kornberg建立了核小体模型。
复制中的核小体
在DNA复制时, 每一个核小体暂时分成两半, 允许DNA聚合酶复进行DNA的复制。在电镜下观察正在复制的染色质时, 可以看到复制叉上有一条DNA链没有核小体结构。并进一步证明, 有核小体的是先导链, 无核小体的是滞后链。
转录中的核小体
通过核小体转录的络纱模型(spooling model)。在该模型中,在DNA转录时, 核小体是全保留的, 通过“DNA成环机制”(DNA looping),使RNA
聚合酶通过核小体
通过核小体,DNA长度压缩了7倍,形成直径为11nm的纤维
螺线管(Solenoids) 染色体三级结构
由核小体组成的直径10nm的纤维螺旋化,每圈6个核小体,形成外径30nm、内径10nm纤维,H1组蛋白位于螺线管的内侧(长度又压缩6倍)。
第三节 染色体的高级组装
一、侧环(Loop)染色体的四级结构
由螺线管中特定区域与细胞核基质结合后收缩形成。侧环长度约63000bp, 相当于一个复制单位。染色体联会及重组可能发生于该级结构水平上。(50圈/环)
二、染色单体纤维(chromatid fiber)染色体的五级结构
以细胞核基质为支撑基础的侧环结构进一步螺旋化,形成18个侧环组成的圆盘。许多圆盘上下重叠形成粗度约为200-300nm、中空的管状结构,内侧为染色体骨架,称染色单体纤维。
第四节 中期染色体
染色单体纤维进一步螺旋化。
主缢痕和次缢痕处未发生这种螺旋化。
5种组蛋白在结构和功能上有什么异同?
? 组蛋白是真核生物染色体的基本结构蛋白, 是一类小分子碱性蛋白质, 有五种类型:H1 、H2A 、H2B 、H3 、H4(表11-4),它们富含带正电荷的碱性氨基酸, 能够同DNA中带负电荷的磷酸基团相互作用。
? 5种组蛋白在功能上分为两组: 一组是核小体组蛋白(nucleosomal histone), 包括H2A、H2B、H3和H4, 这四种组蛋白相对分子质量较小(102~135个氨基酸残基), 它们的作用是将DNA分子盘绕成核小体。它们没有种属及组织特异性, 在进化上十分保守, 特别是H3和H4是所有已知蛋白质中最为保守的。H1属于另一组组蛋白, 它不参加核小体的组建, 在构成核小体时起连接作用, 并赋予染色质以极性。H1有一定的组织和种属特异性。H1的相对分子质量较大, 有215个氨基酸残基, 在进化上也较不保守。
DNA包装成核小体大约压缩了7倍, 请说明计算的依据。
? 一个核小体的直经是10nm, 由200个碱基对的DNA组成, 每个碱基对长度为0.34nm, 一个核小体伸展开来的长度是70nm, 因此, DNA包装成核小体, 大约压缩了7倍。
疖(Knob)
染色粒(chromomere)
部分染色质在细胞分裂前期,尤其是粗线期聚集而成的呈念珠状颗粒,直线排列于染色体上,是DNA与蛋白质结合,在核小体组装染色体过程中形成的一种局部螺旋化结构。
这种组装不是随机的,它随物种、细胞类型、细胞分裂阶段不同而异。因此可作为一种形态标记来识别特定物种的特定染色体。
异染色质的染色粒一般比常染色质的染色粒大,且染色深。 疖(Knob)
特别大染色特别深的染色粒称“疖”。它可作为某些物种特定染色体的识别标志。例如玉米第9、第10染色体。
端粒(Telomeres)
末端特化的着色较深部位。由端粒DNA和端粒蛋白组成。 作用:防止染色体降解、连粘,抑制细胞凋亡,与寿命长短有关。
三、染色体核型和核型分析
1.核型:
指染色体组在有丝分裂中期的表型, 是染色体数目、大小、形态特征的总和。在对染色体进行测量计算的基础上, 进行分组、排队、配对,
并进行形态分析的过程叫核型分析。
动物、植物、真菌等真核生物的某一个体或某一分类群(亚种、种、属等)的细胞内具有的相对恒定特性的单倍或双倍染色体组的表型。染色体的特征以有丝分裂中期最为显著,主要包括染色体的数目、长度、着丝粒的位置、随体(指某些染色体末端的球形小体,由着色浅而狭细的次缢痕与染色体臂相连)与副缢痕的数目、大小、位置,以及异染色质和常染色质在染色体上的分布、染色体分带类型、同位素渗入等。也称“染色体组型”。
将一个染色体组的全部染色体逐条按其特征画下来,再按长短、形态等特征排列起来的图称为核型模式图,它代表一个物种的核型模式。 有丝分裂染色体核型分析所依靠的形态指标:
① 染色体长度
② 着丝粒位置
③ 次级缢痕的有无和位置
④ 随体的有无、形状和大小
⑤ 用低温处理加Feulgen反应染色显现的异染色质区的分布(数量、位置和大小),特别是用Giemsa法染色或荧光染色显现的带型。 核型分析标准:
根据染色体长度、着丝点位置、长短臂比、随体有无等特点进行编号。 粗线期染色体核型分析所依靠的形态指标:
① 染色体长度
② 常、异染色质
③ 染色粒数目和分布
④ 端粒
⑤ 核仁组织区的有无和位置
⑥ 着丝粒位置
2. 核型进化
染色体基数的变化:
基数增加:炮弹果属植物,n=14 着丝粒错分裂 n=15, n=16
基数减少:还阳参属植物,n=6 不等易位 n=5, n=4, n=3 染色体形态的变化:对称核型 非对称核型;随体
染色体大小的变化:单子叶植物染色体〉双子叶
四、染色体分带和带型分析
指借助于某些物理、化学处理,使中期染色体显现出深浅、大小、位置不同的带纹,特定的染色体其带纹数目、位置、宽度及深浅程度都有相
对的恒定性,可以作为识别特定染色体的重要依据。
荧光分带技术:
Giemsa(吉姆萨)分带技术 C-带, Q-带,G-带等
染色带的类型
1. C带: Centromere Heterochromatin or constitutive Heterochromatin带。
主要显示着丝粒、端粒、核仁组织区或染色体臂
上某些部位的组成型异染色质。因期分带部位不同,有着丝粒带、端粒带、核仁组织区带和中间带等相应的名称。
2.N带: Nucleolus Organizer Region(NOR) 带。
核仁组织区的异染色质与其他部位的异染色质有所区别。用N带技术(热磷酸盐处理)可专一地显示核仁组织区。目前植物上利用同一技术也可显示染色体的其他部位,如着丝粒、染色体臂上的某些特殊区带等。
3. G带: Giemsa 带
分布于染色体的全部长度上,以深浅相间的横纹形式出现。现已证明,有丝分裂中期染色体上的G-带、唾腺染色体上的横纹、粗线期染色体上的染色粒,三者在部位上是准确一致的,因此G-带显示的实际上是染色粒。
4. R带: Reverse带, G带的反带。
植物染色体C-带主要包括以下四种带:
1. 着丝粒带(Centromeric Band):
指着丝粒及其附近的带。
2. 中间带(Intercalary Band):
分布在着丝粒至末端之间的带。
3. 末端带(Telomere Band):
位于染色体两臂末端的带。
4. 核仁组织区带(NOR band):
位于NOR区的核仁染色体专一带。
染色体核型分析和带型分析的应用:
1. 物种起源与进化
2. 染色体结构和数目变异
3. 染色体与遗传的关系
4. 染色体标图
5. 染色体工程育种
第四章 染色体的运动
细胞周期:指从一次细胞分裂完成到下一次细胞分裂结束所经历的全过程。
第一节 有丝分裂中的染色体
1. 染色体的复制和有丝分裂的准备
间期(表面上的平静)DNA复制(核小体复制) DNA转录 蛋白质合成
有丝分裂的启动:多个基因的表达与调控
MPF, 成熟促进因子:能够促使染色体凝集,使细胞由G2期进入 M期的因子。
Cdks (cyclin-dependent protein kinases),周期蛋白依赖性蛋白激酶:主要在细胞周期调控中起作用的蛋白激酶,由于受周期
蛋白的激活而得名。真核细胞中主要有三种类型的周期蛋 白依赖性的蛋白激酶。
Cyclin, 细胞分裂周期相关蛋白:参与细胞周期调控的蛋白,并
且其浓度在细胞周期中是浮动的,呈周期性变化。随着细 胞周期阶段的不同,有时浓度高大几千倍,有时有降为 零。周期蛋白作为一种调节亚基,与周期蛋白依赖性的蛋 白激酶结合并将之激活。
2. 有丝分裂的各个主要时期
2.1 前期
a. 染色质浓缩为染色体 b. 核膜解体
c. 核仁消失 d. 形成纺锤体
2.2 中期
2.2.1 前中期
? 染色体赤道板集合
? 着丝粒定向
? 染色体分布
秋水仙素、巯基乙醇和低温可以破坏纺锤体形成
中期 染色体高度螺旋化,适于进行细胞学研究
染色单体间平行排列,秋水仙素处理,染色体臂分 离,着丝粒处相连:
2.2.2 后期 染色体运动速度:0.2~5?m/min
2.2.2 末期及细胞质分裂
染色体到达两极,子核形成,产生2个子细胞
如何利用胸腺嘧啶和秋水仙素获得同步培养的细胞?
高浓度的胸腺嘧啶能够阻断DNA合成所需的核苷酸的合成, 因此将细胞群体培养在具有高浓度的胸腺嘧啶的培养液中时, 非同步化的细胞能够正常地通过细胞周期, 但到达S期时, 因DNA的合成被阻断, 这些细胞不能顺利通过S期进入G2期。经过对S期的短暂阻断, 再改变胸腺嘧啶的浓度, 解除抑制, 所有的细胞都开始DNA的合成, 即获得处于同步生长的细胞。
秋水仙素可抑制微管的聚合, 因而抑制有丝分裂器的形成, 将细胞阻断在有丝分裂的中期,适当时间后解除秋水仙素的作用,即获得处于中期的同步化的细胞。这一方法称为中期阻断法。
第二节 减数分裂中的染色体
减数分裂:是一种特殊方式的细胞分裂,是在配子形成过程中发生的,包括两次连续的核分裂,但染色体只复制一次,因而在形成的四个子细胞核中,每个核只含有单倍数的染色体,即染色体数减少一半. ? 减数分裂包括两次连续的核分裂:
减数分裂I和减数分裂II。染色体只分裂1次。
? 主要事件有:
同源染色体联会、配对、重组和染色体数目减半。
1 细线期
2 偶线期 前
3 粗线期 期
4 双线期 I
5 终变期
6 中期I
7 后期I
8 末期I
减数I 减数间期 9 前期II 10 中期II 11 后期II 12 末期II 减数II 减数分裂前的间期(premeiotic interphase, premeiosis)
G1
S: 较长,合成总DNA的99.7%
G2: 有丝分裂向减数分裂转变,减数分裂组蛋白
2. 前期I:同源染色体配对、重组。
5个亚期(substages):
细线期
偶线期
粗线期
双线期
终变期
2.1 细线期(Leptonema)
(凝集期;花束期): 细长线状,核仁存在。
2.2 偶线期(Zygonema):(配对期或合线期)
同源染色体开始联会(联会复合体),二价体(四分体)。
同源染色体:指分别来自父母双亲、形态和遗传内容都十分相似的染色体。
同源染色体的对应部位相互开始紧密并列,逐渐沿纵向配对在一起,称为联会(synapsis)。
细胞内2n条染色体可配对形成n对染色体。配对的两条同源染色体称为二价体(bivalent)。细胞内二价体(n)的数目就是同源染色体的对数。
2.3 粗线期: (重组期)发生交换, 可见二价体。
2.4 双线期(Diplotene): (合成期) 联会消失, 配对松弛,可见交叉结(Chiasma
2.5 终变期(Diakinesis)(浓缩期) 交叉端化。
染色体超螺旋化,有相互分开的趋势,纺锤体形成
3. 中期I(Metaphase I, MI)
核膜、核仁消失,染色体成对排列于赤道板上,同源染色体的着丝粒取向两极,纺锤丝附着于着丝点上,着丝粒不分裂。
染色体构型(Chromosome configuration)
单价体,二价体,多价体,异型二价体
4. 后期I(Anaphase I, AI)
配对的同源染色体分离,向两极移动,姊妹染色单体不分开。染色体减半 , 2n n
5. 末期I(Telophase I, TI)
核膜、核仁重现。
连续型胞质分裂(Successive Cytokinesis):一些生物在减数分裂I末期,细胞核之间形 成细胞板。同时型胞质分裂(Simultaneous Cytokinesis):另一些生物,第一次分裂后不形成细胞板,胞质分裂被延迟到第二次分裂结束时进行。
6. 减数分裂II
(1)前期II:每条染色体含2条姊妹染色单体,染色体数是n。
(2)中期II:染色体着丝粒排列在赤道板上。
(3)后期II:着丝粒纵裂,姊妹染色单体向两极移动。
(4)末期II:核膜重新形成。
减数分裂的生物学意义何在?
①减数分裂保证了有性生殖生物在世代交替中染色体数目的恒定
雌雄配子的融合, 把不同遗传背景的父母双方的遗传物质混在一起, 其结果既稳定了遗传,又添加了诸多新的遗传变异, 大大增强生物对千变万化环境的适应能力。然而, 如果没有一种机制使精卵细胞染色体数减少一半, 那么精卵细胞的融合, 将使染色体数倍增下去, 细胞的体积也就不断地膨胀, 细胞将不能适应环境而遭淘汰。减数分裂保证了生殖细胞在细胞周期中染色体的单倍化,然后通过受精作用还原为二倍体。没有减数分裂,有性生殖将是不可能的。
②减数分裂是遗传重组的原动力,增加了生物多样性
减数分裂也是遗传变异产生的主要原因。在生物进化过程中,如果没有遗传变异,生物就不能适应环境的变化,就会失去长期生存的能力。 在减数分裂过程中,有两种方式发生遗传重组。一种是通过亲代染色体在单倍体细胞中的自由组合,产生的配子所含的染色体在组成上既有祖父的也有祖母的。第二种方式是同源染色体配对时发生的DNA交换。这种遗传重组过程产生的单个染色体中既有父本的也有母本的基因。减数分裂就是通过这样两种机制产生遗传上独特的四个单倍体细胞,每个细胞都含有新重组的遗传信息。
后期I同源染色体的成员移向两极是随机的,非同源染色体的组合式自由的。
非姊妹染色单体交叉与片断交换
染色体组 genome 指配子中所包含的染色体或基因的总和。H.Winkler(1920)最初提出,单倍性的一整套染色体为一个染色体组。这是最先所给与染色体组的基本概念。木原均(1980)又赋与此概念以功能上的含义,即把各种生物为保持其生活机能协调谐和而不可或缺的一组染色体作为一个染色体组。在一个染色体组为A的二倍体生物中,体细胞与生殖细胞的染色体组分别为AA和A。单倍体的产生证明了仅一套染色体组即能维持生物的生存。在二套染色体中如其所包含的所有染色体彼此两两相同时称为同源染色体组,相反,如所有染色体都不相同,则称异源染色体。处于二者之间则为部分同源染色体。
基因组,Genome,一般的定义是单倍体细胞中的全套染色体为一个基因组,或是单倍体细胞中的全部基因为一个基因组。可是基因组测序的结果发现基因编码序列只占整个基因组序列的很小一部分。因此,基因组应该指单倍体细胞中包括编码序列和非编码序列在内的全部DNA分子。说的更确切些,核基因组是单倍体细胞核内的全部 DNA分子;线粒体基因组则是一个线粒体所包含的全部DNA分子;叶绿体基因组则是一个叶绿体所包含的全部DNA分子。
染色体核型
染色体组型
染色体构型
第三节 远缘杂种的减数分裂染色体行为及染色体组分析(Genome analysis)
1. 染色体配对的基础:
同源性识别
联会前联合
形成SC
2. 杂种的减数分裂行为
多价体:
三价体III
四价体IV
单价体 I
减数分裂中单价体的行为:
1)AI分向两极,MII正常分离,2/4孢子中具有该染色体
2)AI正常分离,AII随机分离
3)AI着丝粒错分裂为等臂染色体,AII为端着丝粒染色体
4) MI不与纺锤丝相连,丢失在细胞质中形成微核
3. 染色体组分析-- 分析异源多倍体的二倍体供体
根据形态学、组织学、解剖学及生理生化方面的资料选择几个二倍体侯选物种作为测验种(Analyser),与待测的多倍体目标物种(Object)杂交,观察杂种F1的染色体配对行为,综合所有杂种的染色体配对资料推导出目标物种的染色体组来源。
12.1.2 细胞周期的研究方法
为了研究细胞周期的不同阶段的生化特性必须获得细胞周期一致性的细胞, 这就是细胞的同步化(synchronization)。
细胞同步化分为自然同步化和人工同步化。
自然同步化是自然界存在的现象, 人工同步化是利用细胞培养的方法, 用各种理化因素处理获得的同步化生长的细胞。常用的细胞人工同步化的方法分为选择同步化、诱导同步化或者两者的结合。
■ 诱导同步法(induction synchrony)
控制培养条件, 将非同步培养中的所有或大部分细胞暂时性地阻止在细胞周期的某个阶段, 最终使所有细胞达到同步化生长。常用的手段有改变温度、添加代谢抑制剂将细胞阻止在细胞周期的某一阶段。
● 胸腺嘧啶阻断技术(thymidine block technique) 高浓度的胸腺嘧啶能够阻断DNA合成所需的核苷酸的合成, 因此将细胞群体培养在具有高浓度的胸腺嘧啶的培养液,可获得同步化的细胞。
● 中期阻断法 某些药物,如秋水仙素可抑制微管的聚合, 因而抑制有丝分裂器的形成, 将细胞阻断在有丝分裂的中期。 如何利用胸腺嘧啶和秋水仙素获得同步培养的细胞?
选择同步法(selection synchrony)
用物理方法将处于细胞周期中同一阶段的细胞从非同步的群体中分离出来。 选择同步化可克服同步化过程中对细胞的毒性影响。常用的方法有:
● 有丝分裂选择法是根据细胞在细胞周期的不同阶段的生理变化设计的一种方法。此法的优点是不受药物的影响, 同步化程度高, 不足之处是分离的细胞少, 手续繁琐。
● 细胞沉降分离法 此法主要用于悬浮培养的细胞,也可用于贴壁生长的细胞, 但获得的细胞的同步化程度有限, 因同一时相的细胞大小并非都是一致的。 有丝分裂选择法和细胞沉降分离法获得同步化细胞方法的原理有何不同?
■ 的利用
利用条件突变体可研究细胞周期中的重要事件,发现细胞周期基因。例如温度敏感突变使细胞对于某种温度敏感,这样可用正常温度培养细胞而用突变温度来研究突变的事件。利用条件突变,在酵母中鉴定了几十种细胞分裂周期(cell-division cycle,cdc))突变体, 发现了一些重要的细胞周期调控基因。
第五章 染色体的功能
染色体是遗传信息(基因)的载体
遗传的染色体学说的直接证明
摩尔根发现伴性遗传,第一次把特定基因(果蝇白眼基因)与特定染色
体(X染色体)联系起来,为遗传的染色体学说提供了第一实验证据 X染色体不分开(nondisjunction)现象
Bridges推测这些例外是由于X染色体不分开造成的,即两条染色体在减数分裂中没有分开,一起进入同一极,产生的卵子是XX或者O。 X染色体的次级不分开现象
次级例外的产生,是由于初级例外的白眼果蝇在减数分裂中X染色体的次级不分开(secondary nondisjunction)造成的。
第一节 连锁与重组
1. 连锁的发现
2. 重组
2.1 相互重组(Reciprocal recombination)即交换(Exchange , Cross-over)
特征:通过遗传重组产生1对互补的重组染色单体。
2.1.2 影响交换频率的生物学和非生物学因素
1)干涉(Interference): 指一个交换事件的发生阻碍(正干涉)或促进(负干涉)另一个交换事件的现象,如果不考虑染色单体,称染色体干涉,如果指前一交换所涉及的染色单体影响相同的两个染色单体邻位上的另一交换,称染色单体干涉。
2)染色体位置效应:在所有测试的生物中着丝粒附近的交换频率明显下降。着丝粒对交换的抑制效应可能与着丝粒区的异染色质结构有关。 “重组热点” “重组冷点”“连锁不平衡”
3)性别:雄性果蝇中通常不发生交换,在雌性家蚕中也未观察到交换;鸽子中雄性交换值总大于雌性,而鼠类中正好相反。
4)基因型:
5)年龄及温度:极端高温和低温有提高交换频率的趋势。辐射对交换影响很大,可刺激果蝇交换。温度和辐射能诱发正常条件下不发生交换区域发生交换。二价离子,如钙离子、镁离子等,能够改变交换的数量。过量的钙离子降低交换频率;在低于正常水平时,交换频率显著增加。另外,年龄等其它外部条件也可能影响某些基因间的重组。
2.2 非相互重组(Non-reciprocal recombination)(Flash)
重组产物不互补,表现为遗传物质的不对称交流,等位基因分离严重偏离孟德尔分离比例。
2.2.1 基因转换(Gene Conversion)
指减数分裂过程中一个等位基因转变为另一等位基因的现象(6 : 2)
2.2.2 减数后分离(Post-meiotic segregation)
由于重组过程中形成的杂合双链DNA分子未能及时进行错配碱基的修复,杂合双链部分在随后的细胞周期中随着DNA的复制而纯合,并随染色单体分裂(5 : 3和异常4 : 4)。
3. 连锁图谱(Linkage Map)或遗传图谱(Genetic Map)
连锁群(linkage group):一条染色体上的所有基因,在传递行为上具有共分离的趋势,共同组成一个连锁群。一个生物连锁群的数目等于其配子染色体数。
遗传图谱: 反映位于同一染色体上的基因间的排列顺序和相对距离,图谱的建立以交换为基础,基因间距是由交换频率转换而来,1%的交换值作为一个作图单位,常称为分摩(Centi-Morgan, cM)。
4. 连锁和重组的生物学意义
? 遗传稳定性:一条染色体即为一个连锁群
基因精确分配
功能和进化需要减少重组
Y染色体
操纵子
交叉保证同源染色体均等分配
? 生物多样性:
无重组: 2n 种基因型(n=配子染色体数)
有重组: 2m 种基因型(m=物种基因总数)
第二节 重组机制
1. 交换的细胞学证据(Creighton and Mclintock,1931)
2. 交叉与交换
交叉是同源染色体交换的结果。
3. 重组发生的时间:减数分裂前期
4. 遗传重组机制
第三节 性别决定与性染色体的功能
1. 性别决定类型
1.1 性染色体与常染色体(sex-chromosome and autosome)
与性别有关的一对形态、大小不同的同源染色体称为性染色体(XY或ZW),除性染色体之外的其他染色体称为常染色体(A)。
1.2 性别决定(sex determination
1.2.1 多数高等植物和低等动物:雌雄同株(体),无性 别决定问题
1.2.2 高等动物和某些植物:多数为雌雄异体(株),性别由性染色体的构成决定,可以分为:XY型性别决定:人类、哺乳类、两栖类、鱼类、
昆虫、植物。如人类:44A+XY ZW型性别决定:鸟类、爬行类、鳞翅目昆虫:如家蚕:54A+ZW XO型性别决定:直翅目昆虫如蝗虫、蟋蟀、蟑螂。
1.2 染色体倍性与性别蜂类和白蚁
1.3 植物的非染色体决定
2. 性别决定机制
2.1 性基因平衡理论(Bridges,1925)
物种有两套不同的基因与雌雄性别的分化有关,一套为雄性的决定基因,位于常染色体上;另一套为雌性决定基因,位于X染色体上,两套基因的平衡决定性别的遗传.
性比(X : A ratio): 决定性别
2A+2X X:A=1 雌性(XX, XXY)
2A+1X X:A=0.5 雄性 (XY, XO)
0.5 <X:A <1 雌雄间性
Y染色体与雄性育性有关,对性别无决定作用。
2.2 Y染色体的雄性决定作用
剪秋箩: XX: 雌性 XY:雄性 Y染色体存在,植株即发育为雄株或雌雄同株 。
X:Y比值决定雄性发育程度:
X:Y<5 雄株
1.5 < X:Y<40 雌雄同株
3. 性染色体的起源进化
4. 剂量补偿与X失活
剂量补偿效应(Dosage compensation): 使细胞中具有两份或两份以上基因的个体和只有一份基因的个体出现相同表型的遗传效应。 巴氏小体( Barr body): 即性染色质。无论一个个体有多少条X染色体,在体细胞中只有1条X染色体具有转录活性,其余的X染色体全部失活沉默,在细胞学上表现为异固缩,两个端粒互相靠近形成功能型异染色质体存在于核膜内侧,这种异染色质称性染色质,19xx年由Barr发现,故称巴氏小体。
5. 性别分化、变异与控制
5.1 性别分化 激素
5.2 性别变异
人类、鼠、家畜:睾丸雌性化综合症
果蝇:TRA基因使XX个体转变为不育的雄蝇
5.3 性别控制
? 改变影响性别分化的环境条件:温度、激素
? 哺乳类动物,早期细胞学和分子遗传学检测:
雌体巴氏小体、雄体SRY序列
? 利用性连锁基因:家蚕
第六章 特殊类型的染色体
? 多线染色体(Polytene chromosome)
? 灯刷染色体(Lampbrush chromosome)
? B染色体( B or Extra chromosome)
? 端着丝粒染色体(Telocentric chromosome
第一节 多线染色体
核内DNA多次复制产生的子染色体平行排列, 且体细胞内同源染色体配对, 紧密结合在一起, 从而阻止了染色体纤维进一步聚缩, 形成体积很大的由多条染色体组成的结构叫多线染色体。
多线化的细胞处于永久间期, 体积也相应增大, 它存在于双翅目昆虫的幼虫组织内, 如唾液腺、气管、前肠、中肠及马氏管细胞中;原生植物;显花植物的反足细胞、胚柄等组织中。
核内有丝分裂(Eudoreduplication):染色体的DNA链经过复制,染色线分离,而细胞并不分裂,形成核内多倍性。
多线性(Polytene):
染色体的DNA链多次复制,而细胞并不分裂,染色线亦不分离,形成多线染色体。
1. 多线染色体的形态特征
1.1 多线性和巨大性
1.2 体细胞配对
? 同源染色体配对是减数分裂期细胞的一个特征性过程,它也可以发生在其它类型的细胞中,这种现象通常被称为体细胞配对。 ? 果蝇唾液腺多线染色体在整个间期内都并行紧靠在一起,是体细胞配对中最明显的类型。
1.3 横纹带
2. 多线染色体的膨突及其功能
膨突(puff or Balbiani ring):指多线染色体上从小到大的隆起部位。
? 在不同的组织中,膨突在染色体上出现的部位不同;
? 同一组织中,在不同的发育时期,多线染色体上膨突出现的部位
也不一样。
? 特定的基因只有在特定的发育阶段才能发挥作用,由此推测膨突可能是基因活动的场所,即基因活跃区。
? 并不是染色体上所有的横纹带都能形成膨突,在一种摇蚊中,估计约2000条带中有15%与膨突有关。膨突的大小也有很大的变化,可以为带纹稍有膨大到向外扩散得很厉害形成得大膨突。
第二节 灯刷染色体
灯刷染色体(Lampbrush chromosome)
为转录活跃的一类巨型染色体,存在于卵母细胞第一次减数分裂前期的双线期,二价染色体配对后,染色丝两侧产生无数突起,呈灯刷状,称灯刷染色体。
第三节 B染色体
又称辅助染色体、超数染色体或额外染色体,指真核生物中常染色体以外的一类染色体。
特点:
? 长度短
? 数目不定
? 全部染色深或在端粒和着丝粒区染色深
? 玉米和黑麦中亚端着丝粒染色体(臂比大)
? B染色体的形态、结构及起源
1.1 形态
短:1/3~2/3 A 染色体
臂比:近端着丝粒(黑麦和玉米)
亚中着丝粒(寄生黄蜂)中着丝粒
1.2 结构
全部或大部分为异染色质与A染色体缺乏或少同源序列
1.3 起源于A染色体。
缪勒棘轮(Mullers Ratchet)
A染色体重排或不分离产生类似于三体性质的初生B染色体,带有与其祖先染色体相同的基因。初生B染色体可缺失部分功能基因,B染色体基因越多,与A染色体重复越多,对个体损害越大。因此初生B染色体与突变的B染色体比较在进化中处于不利地位,一旦丢失,就不能再生,从而使“棘轮”向前转动一格。
2. B染色体的减数分裂行为不与A染色体配对。
1个B染色体:单价体
2个B染色体:二价体 AI正常分离,AII不分离
3个B染色体:多价体、二价体、单价体
3. B染色体的数量分布、变异性及积累的机制
3.1 数量多样性:
不同个体,
不同物种,
不同组织,
不同细胞。
3.2 积累机制:不分离机制(Non-disjunction)4种主要类型:
4. B染色体的遗传学效应
4 .1 对A染色体减数分裂配对的影响对大麦族物种部分同源染色体配对具有调节作用。
4.2 对A染色体总长度的影响使A染色体总长度增加。
4.2 对A染色体基因表达的影响 百合属:酯酶同工酶
4.2 对生活力和育性的影响10-15条:无影响;15-25条:影响生活力、结实性、育性;30-34条:生活力极差、不育
第四节 端着丝粒染色体和等臂染色体 端着丝粒染色体:指具有末端着丝粒的染色体。
等臂染色体:指具有2条完全相同的臂中央着丝粒染 色体。
1. 端体的来源、体细胞稳定性及传递
? 端着丝粒染色体的稳定性取决于其着丝粒的完整性。
? 单端体雌配子传递率约50%,雄配子传递率约4%。
? 端体的应用
3.1 鉴定单体:单价体变迁(Univalent shift )
3.2 鉴定相互易位(第9章)和异源易位(第10章)
3.3 基因定位
单端单体: 同单体定位
单端三体:同三体
双端二体:同缺体
可以将基因定位于特定染色体臂上,根据单端二体、单端体自交后代基因分离与染色体组成分析,可以估算目标基因与着丝粒的相对距离(祥见第八章)
4. 等臂染色体(Isochromosome)
假等臂染色体由1对同源染色体的2条不同臂的末端区段发生相互易位而形成的。其遗传组成上与等臂染色体相似,染色体臂的末端同源,而靠近着丝粒的区段非同源。
4.1 等臂染色体的减数分裂配对行为
4.2 等臂染色体的遗传学效应
1)为缺失-重复染色体
2)非交叉遗传
交叉遗传:为性连锁基因特有的遗传现象,指一个隐性突变基因纯合母本和一个野生型父本杂交后代,雄性像母本,雌性像父本的现象。祖父所具有的某一性状通过女儿传递给孙代。
《细胞遗传学》测验题
1 有丝分裂中期染色体的形态结构特征。
1. 2 染色体的基本结构模型。
1. 3 染色体构型分析有那些方面的应用?
1. 4 构建连锁图谱的细胞学基础是什么?
1. 5 何谓剂量补偿效应?
第七章 染色体结构变异
缺失 (Deletion)重复 (Duplication)倒位 (Inversion)
? 染色体是遗传物质的载体。
? 染色体形态、结构、数目的稳定。
? 细胞分裂时染色体能够进行有规律的传递。
? 染色体稳定是相对的,变异则是绝对的。
? 19xx年发现:电离辐射可以诱导染色体结构变异。
? 引起染色体结构变异的原因──先断后接假设:
? 染色体折断,重接错误,结构变异,新染色体
? 染色体折断是结构变异的前奏。
一、缺失(deletion)-由于染色体臂丢失部分遗传物质所产生的染色体结构变异。
Bridges, 1917 在果蝇中首次发现。
玉米,大麦,小麦,番茄,蝗虫及人类均存在缺失。
1. 缺失的起源 自发或人工诱导
1) 诱变剂处理
物理诱变剂:如X-射线、γ-射线、快中子等
化学诱变剂:如甲基磺酸乙酯(EMS)
? 在物理诱变剂及化学诱变剂作用下,染色体臂发生断裂,如重接
不及时,则无着丝粒的染色体断片会在细胞分裂过程中丢失,而带着丝粒的染色体片段的断头则可能愈合成为稳定的缺失体;或者2个姊妹染色单体的断头融合,形成双着丝粒染色体,在细胞分裂后期称为染色体桥,进行断裂-融合-桥循环。
? 因每个循环中断裂的部位不同,又会进一步产生一系列缺失、重复及环状染色体,这些染色体片段上的基因也随之缺失、重复,在细胞中不规则分离。
2) 不等交换
3) 杀配子染色体效应(gametocidal chromosomes)尾状山羊草,柱穗山羊草 , 高大山羊草
4) 组织培养
(Tissue culture)
5) 单价体错分裂
2. 缺失的类型 中间缺失 顶端缺失 整臂缺失
3. 缺失的细胞学特征
1)无着丝粒断片:这种断片可残存于细胞质中形成微核。由于其它染色体结构变异和数量变异也会导致微核,所以应该进一步调查减数分裂过程。若同时观察到异型二价体或后期II姊妹染色单体长度不等。2)染色体核型、带型变化:缺失发生后,染色体的臂比随之改变,分带后可能会发现部分带纹丢失。
3)染色体配对:在玉米减数分裂粗线期可以清楚观察到同源染色体沿着长轴方向配对。
? 如果染色体发生缺失,在杂合状态下其粗线期将表现不对称配对。 ? 对于顶端缺失,则在配对染色体构型上可呈现一段单体;如果发生中间缺失,则正常染色体在缺失部位突出成环。
4. 缺失的遗传学效应
1) 假显性(Pseudodominance): 如果缺失染色体片段包括某些显性等位基因,当缺失纯合体与纯合隐性个体测交时,F1隐性性状可以像正常地显性性状一样得以表现,这一现象称为假显性。
2) 假等位(False allelism): 指紧密连锁的一些基因,它们 在功能上具有等位基因的性质,但在染色体线性排 列上并不等位,由于不同染色体区段的缺失,似乎 表现为等位基因的关系,这种现象称~。
3) 致死效应
? 当缺失的染色体片段很小,或者缺失的基因不很重要时,缺失一
般不会影响到个体的存活能力。但当缺失片段较大,并携带重要基因 时,缺失纯合体往往不能生存。
? 高等植物对缺失的忍耐能力与其染色体倍性有很大关系,二倍体植物,如玉米、大麦、黑麦对缺失耐性很差,尤其是花粉,即使不完全败育,受精过程也竞争不过正常花粉粒。多倍体植物,如普通小麦、栽培燕麦、烟草等,由于部分同源染色体间的遗传补偿性,对缺失的承受能力较强,不仅染色体部分缺失的个体可以存活,而且连缺失整条染色体的有些缺体都能存活,甚至结实。
4) 微缺失与性状变异
? 在诱变育种实践中产生的许多突变体中,相当一部分是由于较短的DNA片段缺失造成的。但这些很短的微缺失与碱基互换、颠换很难区别。理论上,微缺失造成的突变不能恢复,而碱基变化引起的突变则能产生恢复突变。
5. 缺失作图(Deletion mapping)
? 缺失纯合体缺失区段所载的基因丢失导致表型上的变异,缺失杂合体则会表现假阳性。据此可以把某一基因与染色体的特定区段联系起来,确定其在染色体上的物理位置。
? 若创造覆盖全部染色体的一系列缺失体,就可构建某染色体(臂)或整个基因组的基因图谱,它反映基因在染色体上的实际位置,由于是用细胞学方法构建的,又称细胞学图谱(Cytological map)。
二、重复(duplication)-1种引起染色体部分遗传物质剂量增加的染色体结构变异。
1.重复的类型及起源
1.1 染色体间重复(interchromosomal duplication):重复片段整合到其它染色体上的一种染色体结构变异。
1.2 染色体内重复(intrachromosomal)
? 顺接重复(tandem duplication):
重复片段首尾相连,方向一致。
? 反接重复(reverse tandem duplication):
重复片段首-首或
尾-尾相连,方向相反。
? 置换重复(displaced duplication):
重复片段为其他染色体片段
分开,或位于不同臂甚至不
同的染色体。
1.3 重复的起源
? 染色体断裂-重接
? 染色单体不等交换
? 倒位和易位杂合体中的交换
? 转座因子不精确切离或共转导
2. 重复的细胞学特征
1) 核型、带型变化
果蝇X染色体16A区带纹顺接重复
2) 重复杂合体中的重复环:
重复杂合体
3) 拟等臂染色体配对:置换重复位于同一染色体不同臂上
核型、带型变化:重复不仅会改变染色体核型,更重要的是带纹的增加,例如果蝇的棒眼表型就是X染色体上16A区带纹顺接重复结果。
重复环:在重复杂合体中,未配对的重复区段被挤出形成一个重复环,二价体的整体长度和臂比没有改变,这一点与缺失环不同。
拟等臂染色体配对:如果置换重复发生在同一染色体的另一臂上,在减数分裂配对行为上与等臂染色体相似,进行自身配对。
当重复方向相同时,同一染色单体2个臂交换后产生1个环状染色体和着丝粒的断片,不同的染色单体交换后将形成双着丝粒染色体,开始进行桥-断裂-融合-桥循环。
染色单体桥及断片:反接重复杂合体减数分裂配对交换后形成染色单体后期I桥和断片,这与臂内倒位的情况非常相似,不同的是粗线期以前染色体配对二价体中有突出的重复环。
3. 重复的遗传学效应
重复也会引起遗传上的不平衡。但重复一般不会影响个体生活力。无论是纯合还是杂合重复均能较好地生长发育,含有重复染色体片段的配子败育的情况较少。
重复的遗传学效应因重复片段的大小及所携基因的性质不同而异。
3.1 剂量效应(Dosage effect):
? 由于剂量增加,重复的基因在行为上与其在正常野生状态下有所不同的现象,称剂量效应。
3.2 位置效应 (Position effect):在基因剂量相同的情况下,由于基因在染色体上位置不同而引起表型变化的现象。
4. 重复的进化意义
1)物种进化
原核生物 生物进化 真核生物
2)物种形
? 随着原核生物向真核生物进化,其基因组(DNA含量)增大,并产生大量的重复DNA序列 。这是由于遗传物质部分重复所导致的2个相关现象。基因组中一部分DNA经过多次重复形成多拷贝基因族,如核糖体RNA基因、组蛋白基因,或者无编码功能重复序列,如卫星DNA、端粒DNA等,另一些DNA重复后经过突变发生多样化,有可能执行新的功能。所以在一定意义上,重复是生物进化的物质基础,是真核生物遗传结构和功能复杂化的物质保证,是一种重要的生物变异方式。
? 另一些物种的形成过程中染色体区段的重复也发挥重要作用。如玉米中就有大量的染色体内和染色体间重复。这表明玉米起源经历过一个多倍体化及随后的染色体普遍重排过程。
第九章 染色体的异常细胞遗传学行为
第1节 体细胞联会和体细胞交换
1. 体细胞联会(Somatic pairing)体细胞中同源染色体配对的现象。
2. 体细胞交换(somatic crossing-over,mitotic crossing-over) 亦称有丝分裂交换,指体细胞在有丝分裂时发生的染色体交换。以体细胞联会为基础,发生于染色体四线期。可由
DNA合成抑制剂或DNA修复中的特定事件引发,导致隐性有害 基因的纯合。
3. 姊妹染色单体互换(Sister Chromatid Exchange, SCE)
体细胞中同一染色体的姊妹染色单体之间发生互换的现象。 对于分裂的细胞,如将5溴脱氧尿嘧啶核苷(5-BrdU)加入合成DNA的原料中,经过两个分裂周期后,两条染色单体,其中一条DNA链的双股内的胸腺嘧啶核苷均被BrdU取代,另一条只有一股被取代。
用染色剂如吉姆萨和光处理,使双股含BrdU的染色单体着色浅淡,而单股含BrdU的染色单体着色深。
计数SCE数,判断受试物对DNA是否有损伤作用。
第2节 染色体消减
在一些远缘体细胞融合体及受精卵的有丝分裂中,其中一方的全部或绝大多数染色体受到排斥逐渐丢失,而只保留另一方的染色体的现象。
第3节 减数分裂联会变异及其遗传控制
1. 类型:不联会(asynapsis, as)联会消失(desynapsis, des)
2. 原因:基因突变
远缘杂种后代不联会
1条(对)染色体的缺失或添加
第四节 不减数配子
不减数配子的形成机制:
1. 雄配子
1) 减数前异常:
孢原细胞核内有丝分裂:
共双倍性(Syndiploidy):指雄性生殖细胞中两个已分开的细胞核在减数分裂前的融合。
2) 减数分裂异常:
减数分裂核再组(meiotic nuclear restitution): 指在减数分裂 I或II失败后,由未减数的染色体而不是由2个减数 的细胞核形成一个细胞核的过程。
细胞质提前分裂:
3)细胞质分裂异常:
2. 雌配子
1) 联会变异体
2)延迟减数分裂
3)减数分裂II消失
4)异常后期
5)末期II细胞质分裂失败
第五节 染色体的选择性传递
? 减数分裂驱动(Meiotic drive)
指通过染色体传递不均等性而影响群体中基因频率的任何机制,主要发生在结构变异杂合体中
? 减数后有丝分裂不均等:功能极
? 受精竞争
野生小麦:杀配子染色体:3C, 4S, 7el2