实验五 酪蛋白的制备及含量测定
一、实验目的
1.学习和掌握紫外吸收法测定蛋白质含量的原理和方法
2.学习分光光度计的原理和操作方法。
二、实验原理
牛乳中主要的蛋白质是酪蛋白,含量约为35g/L。酪蛋白是一些含磷蛋白质的混合物,等电点为4.7。利用等电点时溶解度最低的原理,将牛乳的pH调成4.7时,酪蛋白就沉淀出来。用乙醇洗涤沉淀物,除去脂类杂质后便可得到纯的酪蛋白。
蛋白质分子中含有酪氨酸、色氨酸及苯丙氨酸残基,它们的结构中具有共轭双链,对紫外光有吸收作用,其最大值在280nm波长处。在此波长附近,蛋白质溶液的光吸收值与其含量(范围是0.1~1.0mg/ml)成正比,因此,280nm的吸光度可用作蛋白质的定量测定。但不同种的蛋白质对280nm波长的光吸收强度因芳香性氨基酸残基含量的不同而有差异,因此需用同种蛋白质作对照,结果才可靠。在半定量测定和纯蛋白的定量测定时,可用280nm的光吸收法。
在分光光度计中,将不同波长的光连续地照射到一定浓度的样品溶液时,便可得到与众不同波长相对应的吸收强度。如以波长(λ)为横坐标,吸收强度(A)为纵坐标,就可绘出该物质的吸收光谱曲线。利用该曲线进行物质定性、定量的分析方法,称为分光光度法。
紫外吸收法操作简便快捷,不消耗样品,低浓度的盐类不干扰测定,在蛋白质和酶的生化制备及其它研究中应用广泛。
三、实验仪器及试剂
95%乙醇(配100mL)、乙醇-乙醚混合液(V:V=1:1)(配200mL)
0.2mol/L NaOH (配250mL)
0.2M pH4.7 HAC-NaAC 缓冲液(配300mL): A液:称NaAc·3H2O 54.44克,定容至2L。 B液:称醋酸 12克,定容至1L。 取A液1770mL,B液1230mL混合即得pH4.7的醋酸-醋酸钠缓冲液3L。
1mg/ml标准酪蛋白溶液:称取酪蛋白1克于研钵中,先用少量蒸馏水湿润后,慢慢加入0.2mol/L NaOH 4ml,充分研磨,用蒸馏水洗入100ml容量瓶中,放入水浴中煮沸15分钟,溶解后冷却,定容至100ml,保存于冰箱内。用时配制成浓度为1mg/ml的溶液。
牛奶、无水乙醚、离心机、天平、滤纸(Φ=9cm)、表面皿、抽真空泵、抽滤装置(布氏漏斗,抽滤瓶,圆形抽滤管,橡皮塞)、40℃水浴、通风柜、紫外分光光度计、试管
四、实验步骤
1、等电点法制备酪蛋白
将20mL pH4.7的醋酸-醋酸钠缓冲液预热至40℃,将20mL牛奶加热至40℃,(牛奶和缓冲液预热的目的?)在搅拌下缓慢地加入20mL预热的pH4.7的醋酸-醋酸钠缓冲液,用精密pH试纸调pH至4.7(如果pH没有调至4.7,对得率有什么影响?),可见溶液变为乳白色悬浮液。待悬浮液冷却至室温,4000rpm离心8min,弃上清,得酪蛋白粗制品。
用6mL水洗脱沉淀1次(酪蛋白溶于水吗?),4000rpm离心5min,弃上清在沉淀中加入6mL 95%乙醇(乙醇作用?),搅拌片刻,将全部悬浊液转移至布氏漏斗(什么作用?)中抽滤,用乙醇-乙醚混合液洗涤沉淀1次,最后用乙醚洗涤沉淀1次(乙醚什么作用?在什么环境中使用?),抽干。将沉淀摊开在干净玻璃或培养皿上,风干,得酪蛋白纯品,准确称量,计算含量和得率。(注意含量和得率的区别)
含量(g%)=(酪蛋白g/100mL牛奶)×100%
得率=(测得含量/理论含量 )×100%,理论含量为3.5g/100mL牛乳
2、紫外吸收法测蛋白质含量(这步和上步的关系?上步中制得的酪蛋白是纯品吗?紫外吸收法测得的蛋白质含量准确吗?)
(1)标准曲线的绘制(标准曲线的横纵坐标的单位是什么?1-8号管的横坐标对应的数值是?表中1-8号管的蛋白质浓度数值是怎么得来的?)
取8支试管,编号后按下表加入试剂。(几号管是用来作对照调零的?)
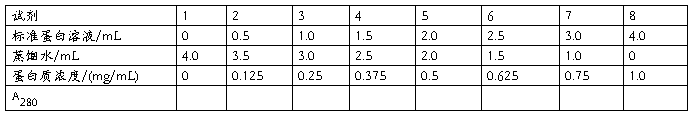
加完混匀后,用紫外分光光度计测A280,以吸光度为纵坐标,蛋白质浓度为横坐标作图。
(2)酪蛋白样品液制备测定
配制酪蛋白含量在0.05~1.25mg/mL(为什么配成这个浓度?怎么配成这个浓度的溶液?)的待测样液进行吸光值测定。为使酪蛋白溶解,可向样品中加入少量0.2mol/L NaOH,再用蒸馏水洗入100ml容量瓶中定容。
取待测酪蛋白溶液1.0mL置试管中,加入蒸馏水3ml,混匀,(以什么为对照调零?)按上述方法测定280nm波长处的光密度值,并从标准曲线上查出待测酪蛋白的浓度。
五、实验结果
1、计算酪蛋白含量和得率,并指出实验步骤中会影响得率高低的具体操作。
2、根据紫外吸收法测出1.0mL样液中酪蛋白的浓度,计算出20mL牛奶中含有的酪蛋白质量。并进一步计算出根据紫外吸收法得到的酪蛋白含量和得率。
3、比较等电点法制备酪蛋白质量和紫外吸收法计算出的酪蛋白质量,得出结论。
第二篇:生物化学资料
生物化学资料
食物中摄入的生物大分子在进入细胞进行分解代谢之前有一个消化和吸收的过程。在各种水解酶的作用下,蛋白质、脂质、多糖和核酸被降解成各自的组成单位,再通过吸收或运输进入细胞被进一步分解。
绝大多数动物消化道缺乏水解β-1,4糖苷键的酶,因此纤维素就无法被水解利用。而带有α-1,4糖苷键的淀粉和糖原则可以在消化道分泌的各种α-糖苷酶催化下被最终水解成葡萄糖单位。单糖进入细胞被吸收的过程是运输蛋白介导的,至少有Na+-单糖共运输蛋白系统和不依赖于Na+的易化扩散运输系统参与催化单糖从肠腔进入小肠上皮细胞的过程。
脂在消化过程中需要由胆囊分泌的胆汁酸(盐)进行增溶。在胆汁酸(盐)的帮助下,大脂滴被分散成小的脂滴,此过程被称为乳化,这大大提高了脂水解效率。吸收需要形成胶束结构,当胶束遇到肠细胞的刷状缘时,脂即被吸收。在肠细胞内,被吸收的脂肪和磷脂在内质网和高尔基体重新被合成并与其他脂和载脂蛋白一起形成乳糜微粒,进入乳糜管,最后汇入血流。
消化道内参与蛋白质消化的蛋白酶起初以酶原的形式被分泌,然后被水解激活。经过蛋白酶消化,食物中的蛋白质在小肠内主要水解成寡肽。寡肽在小肠刷状缘上的肽酶催化下进一步被水解成游离的氨基酸和更小的小肽。小肠上皮细胞面向肠腔的细胞膜上至少有四类依赖于Na+的氨基酸运输蛋白,分别运输酸性氨基酸、碱性氨基酸、中性氨基酸和脯氨酸进入胞内。小肠上皮细胞不需要Na+也能有效吸收二肽和三肽,一旦它们进入胞内,很快被细胞质的肽酶水解成自由的氨基酸。
食物中的核酸在消化道中受到核酸酶的作用会被消化成为核苷酸。带有负电荷的核苷酸需要被在回肠上皮细胞上的与膜结合的核苷酸酶和核苷酶进一步水解成能够被吸收的核糖、碱基和磷酸基团。吸收的过程为主动运输。
真菌也能利用细胞外消化和吸收从环境获取营养。实际上真菌分泌各种消化酶,以便将它们周围的蛋白质、脂肪和糖类水解成为可以被其细胞吸收的氨基酸、FFA、甘油和单糖等。
糖酵解几乎发生在所有细胞,通过糖酵解,葡萄糖或者其他单糖在没有氧气的参与下被氧化成为丙酮酸,并产生NADH和少量ATP。
糖酵解在细胞液中进行,共有十步反应,可划为两个阶段,前五步为引发阶段,葡萄糖与两分子ATP反应,通过磷酸化、异构化、再次磷酸化,形成果糖-1,6-二磷酸,每次磷酸化都需要ATP。果糖-1,6-二磷酸再转变为甘油醛-3-磷酸。甘油醛-3-磷酸再进入第二阶段即产能阶段。在甘油醛-3-磷酸脱氢酶作用下,甘油醛-3-磷酸经历氧化和磷酸化生成甘油酸-1,3-二磷酸和NADH。在这步反应中,砷酸能代替磷酸参与反应,生成不稳定化合物,导致氧化和底物水平磷酸化的解偶联。在磷酸甘油酸激酶的作用下,甘油酸-1,3-二磷酸的高能磷酸基团转移到ADP分子上生成一分子ATP,同时生成一分子3-磷酸甘油酸。3-磷酸甘油酸在磷酸甘油酸变位酶的催化下,转变为2-磷酸甘油酸。后者由烯醇化酶催化形成PEP。在丙酮酸激酶的作用下,PEP将高能磷酸基团转移给ADP形成另一分子ATP和丙酮酸。
在糖酵解过程中,第一步、第三步和第十步为不可逆反应。催化这三步反应的酶是己糖激酶、磷酸果糖激酶和丙酮酸激酶。这三种激酶在催化过程中,表现出典型的“诱导契合”效应。磷酸果糖激酶是哺乳动物糖酵解途径的主要调节酶。它的活性受到高浓度ATP和柠檬酸的抑制,果糖-2,6-二磷酸和AMP的激活。己糖激酶受葡糖-6-磷酸抑制。丙酮酸激酶受高浓度ATP和Ala的抑制,由果糖-1,6-二磷酸激活。血液中的葡萄 1
糖水平下降能引起丙酮酸激酶的磷酸化,导致其活性降低。
糖酵解第一阶段,每一个葡萄糖分子消耗2分子ATP,在第二阶段,每分子甘油醛-3-磷酸产生两分子ATP,因此,通过糖酵解,一分子葡萄糖净产生两分子ATP。
在无氧条件下,为使甘油醛-3-磷酸继续氧化,必须NADH重新氧化成NAD+。在动物和某些微生物体内,乳酸脱氢酶催化丙酮酸生成乳酸,而某些微生物体则将丙酮酸转变为乙醇和CO2。
在有氧条件下,丙酮酸可进入线粒体被丙酮酸脱氢酶系氧化脱羧成为乙酰CoA,NADH则可以通过线粒体内膜上的穿梭系统进入呼吸链氧化放能。
除葡萄糖外,其他六碳糖,如果糖,半乳糖,甘露糖都是通过转变为糖酵解途径的中间产物进入酵解途径。
有氧条件下,葡萄糖的分解并不停止在丙酮酸,而是继续进行有氧分解,最后生成CO2和水,它所经历的途径分为两个阶段,分别为TCA循环和氧化磷酸化。TCA循环是糖、脂肪酸和氨基酸最后分解的共同途径,这一循环的中间体还可作为许多生物合成的前体。
TCA循环在线粒体中进行。丙酮酸进入TCA循环前需通过四步反应先转变为乙酰辅酶A。参加TCA循环的酶共有9种。柠檬酸合酶催化乙酰辅酶A与草酰乙酸反应生成柠檬酸,该酶依次与底物草酰乙酸、乙酰辅酶A和水结合,前后发生两次构象变化,是一个有序的动态催化反应。它是TCA循环中的限速酶,活性受ATP、NADH、琥珀酰辅酶A和脂酰CoA等的抑制。氟乙酰CoA可在它的催化下与草酰乙酸反应生成氟代柠檬酸,而氟代柠檬酸抑制下一步反应。顺乌头酸酶催化柠檬酸与异柠檬酸的互变。异柠檬酸脱氢酶催化异柠檬酸生成α-酮戊二酸,它也是一个变构调节酶,受NAD+和ADP激活,NADH和ATP抑制。α-酮戊二酸脱氢酶复合体与丙酮酸脱氢酶复合体类似,受ATP、产物琥珀酰辅酶A和NADH的抑制,但不受磷酸化与去磷酸化的调节。琥珀酰辅酶A合成酶催化琥珀酰辅酶A和GDP或ADP及Pi发生底物水平磷酸化产生一个GTP或ATP和琥珀酸。琥珀酸脱氢酶是TCA循环中唯一的一个存在于线粒体内膜的酶,催化琥珀酸脱氢生成延胡索酸和FADH2,丙二酸是琥珀酸脱氢酶的竞争性抑制剂。延胡索酸酶催化延胡索酸水合生成L-苹果酸。L-苹果酸和NAD+在苹果酸脱氢酶作用下,生成草酰乙酸和NADH,完成了一次循环。
TCA循环是生物体主要的分解代谢途径,能产生更多的能量货币,其中间产物有的可作为很多合成代谢的前体来源,有的可作为别构效应物调节其他代谢途径。TCA循环还能产生CO2,调节体液的酸碱平衡。
乙醛酸循环只存在于植物和微生物中,此循环可导致偶数脂肪酸净转变为葡萄糖。 磷酸戊糖途径是葡萄糖分解的另一条重要途径,发生在细胞液,一般可将其全部反应划分为氧化相和非氧化相。
氧化相包括三步反应,葡糖-6-磷酸和NADP+在葡糖-6-磷酸脱氢酶的作用下生成内酯和NADPH,这一步是戊糖磷酸途径的调控步骤,葡糖6磷酸脱氢酶受到产物NADPH的竞争性抑制。内酯被内酯酶水解为葡糖-6-磷酸。 后者与另一分子NADP+在葡糖-6-磷酸脱氢酶作用下,生成核酮糖-5-磷酸和NADPH。
非氧化相由5步非氧化的可逆反应组成,由异构酶、转酮酶和转醛酶催化,转酮酶需要TPP作为辅酶,反应的性质是异构或分子重排。通过此阶段的反应,6分子戊糖转化成5分子己糖。非氧化相存在的目的是将戊糖-5-磷酸转变为糖酵解的中间物(果糖-6-磷酸或甘油醛-3-磷酸),从而将戊糖磷酸途径与糖酵解途径联系起来。
2
根据细胞对NADPH、核糖和ATP的需要不同,磷酸戊糖途径能以四种不同的模式存在。
磷酸戊糖途径的主要生理功能是产生NADPH,提供生物合成的还原剂,以及产生5-磷酸核糖作为核苷酸合成的前体。
糖异生指以非糖类物质作为前体净合成葡萄糖。非糖物质包括乳酸、丙酮酸、丙酸、甘油、某些氨基酸和所有TCA循环中间产物等,但不包括偶数脂肪酸。
糖异生途径利用糖酵解的所有逆反应,但需要使用不同的酶克服糖酵解途径中的三步不可逆反应。以丙酮酸为例,它的糖异生首先需要克服糖酵解的最后一步不可逆反应:丙酮酸在丙酮酸羧化酶催化下,消耗一个ATP分子形成OAA;接着,OAA在磷酸烯醇式丙酮酸羧激酶催化下,形成PEP,消耗一分子GTP。形成的PEP可以逆糖酵解而上,直到形成果糖-1,6-二磷酸。在果糖-1,6-二磷酸酶催化下,果糖-1,6-二磷酸C1位的磷酸被水解,产生果糖-6-磷酸,又克服了磷酸果糖激酶催化的不可逆反应。果糖-6-磷酸经逆反应变成葡糖-6-磷酸,后者在葡糖-6-磷酸酶催化下水解成葡萄糖,再次克服了己糖激酶催化的不可逆反应。葡糖-6-磷酸酶位于在肝和肾光面内质网,葡糖-6-磷酸必须先转移到内质网腔才能被水解。
糖异生和糖酵解相互协调,一条途径活跃时,另一条途径就受到抑制。 磷酸果糖激酶和果糖-1,6-二磷酸酶是起调控作用的关键酶。调节的手段主要是别构调节。通常作为磷酸果糖激酶的负别构效应物正好是果糖-1,6-二磷酸酶的正别构效应物,反之亦然。
在激烈运动时,肌肉中酵解产生的丙酮酸转变为乳酸以使NAD+再生,乳酸随血液进入肝脏,通过糖异生转变为葡萄糖,又随血液供应肌肉和脑,这个循环称为Cori循环。
光合作用是指生物体利用光能合成有机分子的过程。能够进行光合作用的生物体被称为光合有机体,它们包括绿色植物、藻类、蓝细菌和某些细菌。光合作用不仅作为CO2固定的主要手段,创造了糖类,为生物体提供了食物,而且还是大气中O2的主要来源。
整个光合作用由光反应和暗反应组成。在光反应中,光能被色素捕获并用来直接驱动H2O的氧化,先后有O2的释放、NADP+的还原和ATP的合成;暗反应利用光反应中产生的NADPH和ATP,通过卡尔文循环将CO2同化成糖类。
光反应包括光能的吸收、传递、转化、水的光解、电子传递、NADP+还原和光合磷酸化等反应。光反应在膜上发生,在光合原核细胞中光合膜充满细胞内,在光合真核细胞中光合膜位于叶绿体内的类囊体膜上。吸收光能的色素分为两类:中心色素(特化的叶绿素a)和辅助色素(藻胆素、类胡萝卜素和除特化的叶绿素a以外的其他叶绿素)。辅助色素收集的光能可以传给中心色素,一个光合单位可简单看为是由几百个集光的叶绿素分子和其他色素分子组成的一个天线系统加上具有光化学反应性的中心色素。
光合作用所吸收的光能处在可见光光谱范围,被吸收的光量子有4种去处:以热形式损失、荧光损失、共振能传递和能量转换,在光合系统中主要利用后两种方式。
光合细菌只有一个光系统,而高等植物和藻类具有两个光系统:PSⅠ和PSⅡ。P700是光系统Ⅰ的中心色素,P680是光系统Ⅱ的中心色素,它们都是处于特化蛋白质复合体中的叶绿素a分子二聚体。PSⅠ和PSⅡ通过在类囊体膜上的空间分离来防止激子失窃。
高等植物参与光反应的各成分在类囊体膜上组装成5种复合体,复合体之间存在流动的电子传递体,这与线粒体内膜上的呼吸链很相似。它们包括光系统Ⅰ、光系统Ⅱ、细胞色素b6/f复合体、铁氧还蛋白NADP+还原酶和CF1F0-ATP合酶。前四种复合体与光能的吸收、传递、转化和电子传递以及跨类囊体膜质子梯度的建立和NADP+还原有关,最 3
后一种复合体是利用质子梯度催化光合磷酸化的酶,它与线粒体内膜上的F1F0-ATP合酶在结构和功能上极为相似。光合磷酸化机制与线粒体中的氧化磷酸化相似。
在C3植物中CO2的固定是由核酮糖-1,5-二磷酸羧化酶催化的,此酶兼有加氧酶的活性。反应发生在叶绿体基质中,第一阶段是CO2固定,产生2分子3-磷酸甘油酸,第二阶段是3-磷酸甘油酸还原为甘油醛-3-磷酸,第三阶段是核酮糖-1,5-二磷酸的再生。
C4植物中CO2的固定涉及两种细胞,叶肉细胞在叶片表面吸收CO2,并用它羧化磷酸烯醇丙酮酸生成草酰乙酸,然后通过转变为苹果酸或天冬氨酸被转运到维管束鞘细胞,在这里脱羧生成产物丙酮酸,产生的CO2进入与C3植物中完全相同的途径被固定。光合作用中的光呼吸会消耗O2和部分光合作用固定的碳,降低光合作用的效率,C4植物通过上述方式,阻止氧气和核酮糖-1,5-二磷酸羧化酶接触,从而降低了光呼吸,提高了光合效率。在景天科植物中存在一种景天酸代谢,它们在夜间吸收CO2生成苹果酸后储藏在液泡中,至白天释放,脱羧产生CO2和丙酮酸,CO2的浓缩和糖类的合成发生在同一细胞、不同时间。
糖原是动物体内葡萄糖的主要储存形式,其降解需要糖原磷酸化酶、糖原脱支酶和磷酸葡糖变位酶。糖原磷酸化酶以磷酸吡哆醛作为辅基,从糖原分子的非还原性末端依次断下一个葡萄糖单位,直到糖原分支点前4个葡萄糖残基处。糖原脱支酶先将分支前以α-1,4-连接的三个葡萄糖残基转移到另一个分支的非还原性末端的葡萄糖残基上或核心链上,接着,它的另一种催化作用将暴露出的α-1,6-糖苷键分解。最后磷酸葡糖变位酶将磷酸化酶的产物葡糖-1-磷酸转变为葡糖-6-磷酸。在肝脏中,葡糖-6-磷酸由磷酸酶转变为葡萄糖进入血液。
糖原合成需要UDP-葡糖焦磷酸化酶、糖原合酶和糖原分支酶。UDP-葡糖焦磷酸化酶催化葡糖-1-磷酸和UTP反应,生成UDP-Glc和焦磷酸。糖原合酶催化UDP-Glc与糖原的非还原性末端葡萄糖残基形成α-1,4-糖苷键,它的催化需要糖原素或者核心糖原为引物。分支酶将糖原链上以α-1,4-糖苷键相连的7个葡萄糖单位一齐转移到同一个或另一糖原分子上新的位置以α-1,6-糖苷键相连形成分支。
磷酸化酶和合酶分别是糖原分解和合成的限速酶。两种酶均可以通过可逆的磷酸化方式进行调节,磷酸化酶的磷酸化形式有活性,去磷酸化形式无活性,而合酶正好相反。AMP对磷酸化酶有别构调控作用,磷酸化酶a和磷酸化酶b都R和T两种状态,当AMP浓度很高时,磷酸化酶b由于与AMP结合,构象由T态转为R态,通过进一步磷酸化转变为磷酸化酶a,使酶完全处于活化的R态。此外,肌肉细胞中的磷酸化酶还受到Ca2+的激活。
葡萄糖对肝脏中糖原代谢起着特殊的调控作用,肝脏中的磷酸化酶a是葡萄糖浓度的感受器,葡萄糖与磷酸化酶a结合后,使磷酸化酶a从R态转变为T态,而T态的磷酸化酶更容易发生去磷酸化反应。
糖原累积症种类很多,主要是由于先天缺少糖原代谢中的某种酶。
脂肪酸的氧化分为α- 氧化、β- 氧化和ω- 氧化,其中β- 氧化是主要的方式。 脂肪酸的β- 氧化由六步反应组成:脂肪酸的活化、脂酰CoA的转运、脱氢、加水、再脱氢和硫脂解。脂肪酸的活化形式为脂酰CoA,催化活化反应的酶是脂酰CoA合成酶。每激活一分子游离的脂肪酸需要消耗两分子ATP。
长链的脂酰CoA不能直接透过线粒体内膜,需要专门的转运系统将细胞液的脂酰CoA转变为基质内的脂酰CoA,其成员包括:CPT-I、转位酶和CPT-Ⅱ。CPT-I位于线粒体外膜,负责催化将进入膜间隙的脂酰CoA上的脂酰基被转移到肉碱的β-羟基上,形 4
成脂酰-肉碱,而游离出来的CoA返回到细胞液。随后脂酰-肉碱在内膜上转位酶帮助下,进入基质并在位于内膜基质一侧的CPT-Ⅱ的催化下将脂酰基转移到基质CoA的巯基,重新生成脂酰CoA,而肉碱又经转位酶反向运输出基质。
脂酰CoA在线粒体基质不断发生脱氢、加水、再脱氢和硫脂解反应,直到其碳链完全氧化裂解成乙酰CoA。在反应中形成的NADH和FADH2进入呼吸链,而乙酰CoA进入TCA循环进一步氧化分解。1分子软脂酸经过7轮β- 氧化循环可以产生106分子ATP。奇数脂肪酸的先是偶数脂肪酸一样进行β- 氧化直到丙酰CoA出现为止。丙酰CoA先后在丙酰CoA羧化酶、甲基丙二酸单酰CoA差向异构酶或甲基丙二酸单酰CoA变位酶的催化下,最终转变为琥珀酰CoA,后者可进入TCA循环被进一步氧化分解。
不饱和脂肪酸一开始照常进行β- 氧化,只是当顺式的双键进入β位以后,需要烯脂酰CoA异构酶改变双键的位置和性质,才能继续进行β- 氧化。如果是多不饱和脂肪酸,还需要2,4-二烯脂酰CoA还原酶的帮助。
大于18个碳原子的脂酰CoA可以进入过氧化物酶体或乙醛酸循环体进行β- 氧化。与线粒体内发生的β- 氧化不同的是,催化第一步反应的酶是脂酰CoA氧化酶。该酶将脂酰CoA失去的电子经过FAD交给O2,形成H2O2,不产生ATP。当碳链被缩短到6~8个碳原子以后,需要进入线粒体才能继续进行β- 氧化。乙醛酸循环体内的β- 氧化酶系可将脂酰CoA完全分解为乙酰CoA。
脂肪酸的α- 氧化是指发生在α- 碳原子上的氧化,像植烷酸这样的脂肪酸因它的β- 碳原子被甲基封闭住而难以直接进行β- 氧化,必须先通过α- 氧化去除1个碳原子以后才可以进行β- 氧化。α- 氧化直接在游离的脂肪酸上进行,它并不需要激活,不产生ATP,既可以发生在内质网,也可以发生在线粒体或过氧化物酶体。脂肪酸的ω- 氧化发生在它的ω- 碳原子上,被氧化的脂肪酸不需要活化,参与ω- 氧化的酶主要位于内质网上,为混合功能加氧酶。
在饥饿、禁食或某些病理状态下,脂肪酸在肝细胞的线粒体基质可转变成酮体——丙酮、乙酰乙酸和D-β-羟丁酸。参与酮体合成的酶有硫解酶、β- 羟-β- 甲基戊二酸单酰CoA合酶、HMG-CoA裂合酶和D-β- 羟丁酸脱氢酶。酮体产生以后离开肝细胞进入血液,随着血液循环到达肝外组织,其中丙酮经肺呼出体外,而另外两种酮体可重新转变为乙酰CoA并进入三羧酸循环氧化放能。肝细胞因为缺乏β- 酮酰CoA转移酶或乙酰乙酸硫激酶,所以不能将酮体再转变为乙酰CoA。如果酮体产生过多将会引起酮症,甚至出现酸中毒。
脂肪酸合成与β- 氧化在反应的场所、酶学、电子受体或供体、脂酰基的载体或二碳单位的形式等方面都具有显著的差别。
脂肪酸合成的前体为乙酰CoA。真核细胞生成乙酰CoA的反应主要发生在线粒体基质,乙酰CoA需要先在基质内与草酰乙酸缩合成柠檬酸,再经内膜进入细胞液。一旦柠檬酸进入细胞液,其中的柠檬酸裂合酶催化它裂解成乙酰CoA和草酰乙酸,草酰乙酸可以经过苹果酸和丙酮酸中间物重新转变为基质内的草酰乙酸。
乙酰CoA的活化由乙酰CoA羧化酶催化。原核细胞的乙酰CoA羧化酶由三个亚基组成,分别具有生物素载体、生物素羧化酶和转羧基酶的功能;真核细胞的乙酰CoA羧化酶是一条多肽链同时具有三种功能。
参与脂肪酸合成的蛋白质或酶通称为脂肪酸合酶,它包括:脂酰基载体蛋白;乙酰CoA:ACP转脂酰基酶;丙二酸单酰CoA:ACP转脂酰基酶;β- 酮脂酰ACP合酶;β- 酮脂酰ACP还原酶;β- 羟-ACP脱水酶;烯脂酰ACP还原酶;硫酯酶。
5
细菌和植物体内的脂肪酸合酶为多酶复合体;哺乳动物的脂肪酸合酶为多功能酶,由两个相同的亚基头尾相连,每一个亚基同时具有7个酶活性和ACP的功能。ACP在脂肪酸合成中的功能是与CoA相似作为脂酰基的载体。
脂肪酸合酶催化的一个二碳单位的延伸共有六步反应组成:引发反应、活化的“二碳单位”的装载、缩合、还原、脱水和再还原。以后每增加一个二碳单位并循环一次,直到形成软脂酸。
机体内其他脂肪酸主要是通过各种修饰反应产生的,修饰反应有延伸和去饱和,分别由延伸酶和去饱和酶催化。动物细胞去饱和能力的不足才导致某些脂肪酸成为必需脂肪酸。
奇数脂肪酸既可以通过α- 氧化从偶数脂肪酸转变而成,还可以直接合成。但直接合成的引物是丙酰CoA。
脂肪酸分解代谢的主要调控位点在脂酰CoA跨线粒体内膜进入基质,受到调节的限速酶是CPT-I,丙二酸单酰CoA能够抑制该酶的活性。
脂肪酸合成的限速酶为乙酰CoA羧化酶,其活性的调节方式有两种:一种由别构调节引起的单体和多聚体形式的互变,多聚体才有活性,另外一种方式是“可逆的蛋白质磷酸化”。乙酰CoA羧化酶的磷酸化形式无活性,去磷酸化形式有活性。促进磷酸化的激酶有PKA和一种直接受AMP激活的激酶,而促进乙酰CoA羧化酶去磷酸化的酶是磷蛋白磷酸酶-2A。
脂肪酸合成的调节还可以在转录的水平上对脂肪酸合酶的量进行调控。
哺乳动物合成胆固醇最活跃的细胞是肝细胞,合成胆固醇的一部分反应发生在细胞液,另一部分在内质网上进行。胆固醇合成的前体是乙酰CoA,需要经过四个阶段的反应:3个乙酰CoA→甲羟戊酸、甲羟戊酸→活化的异戊二烯、6个活化的异戊二烯单位→鲨烯,以及鲨烯→胆固醇。
胆固醇在机体内的运输需要脂蛋白的帮助。血浆中的脂蛋白根据密度从低到高的顺序可分为CM、VLDL、IDL、LDL和HDL。CM由小肠上皮细胞装配和分泌,其主要功能是将食物中被小肠吸收的脂肪运输到脂肪组织和其他体细胞作为燃料,同时将食物中的胆固醇转移到肝细胞;VLDL在肝细胞装配,其功能是将内源的脂肪从肝细胞运输到肝外组织作为燃料,其中的胆固醇和胆固醇酯去向则由LDL决定;IDL由VLDL转变而来,血液中的IDL约有一半被肝细胞吸收,另一半在丢失更多的脂肪后转变成LDL;LDL通过受体介导的内吞被肝外细胞吸收,其主要功能是将肝细胞中的胆固醇转运给肝外细胞,以满足肝外细胞对胆固醇的需求;HDL由肝细胞和小肠上皮细胞装配和分泌,HDL富含各种载脂蛋白,它既可以为其他脂蛋白提供载脂蛋白,也可以从其他脂蛋白接受载脂蛋白。HDL的主要生理功能是参与将肝外细胞多余的胆固醇运回肝细胞的逆向运输。
LDL之中的胆固醇通常被称为“坏胆固醇”,因为它与动脉粥样硬化的发生有关。HDL得到胆固醇以后,既可以转移一部分给正在循环的VLDL,也可以一起被肝细胞吸收,进入肝细胞的胆固醇可被降解成胆汁酸排出体外,也可以重新装配在VLDL,进入循环,在肝外组织重新分配,因此HDL上面的胆固醇被称为“好胆固醇”。
机体内的游离胆固醇可进一步发生代谢转变,既可以酯化成胆固醇酯,也可以作为胆汁酸、维生素D、固醇类激素等物质的前体。催化胆固醇酯化反应有ACAT和LCAT,前者位于胞内,后者位于血液。
胆固醇合成的限速酶是HMG-CoA还原酶,该酶为胆固醇代谢调控的主要位点,调节的方式包括“蛋白质的可逆磷酸化”、酶的降解和酶基因的表达调控。
6
HMG-CoA还原酶具有磷酸化和去磷酸化两种形式,其中去磷酸化形式具有活性。调节磷酸化和去磷酸化反应的酶分别是AMP激活的蛋白激酶和HMG-CoA还原酶磷酸酶。
过多的胆固醇、被氧化的胆固醇衍生物和胆固醇合成途径中的许多中间物能够刺激HMG-CoA还原酶的降解,其中的机制与还原酶本身的结构有关系。HMG-CoA还原酶的跨膜区域含有一种固醇感受结构域,这种结构域能够直接感受到细胞内胆固醇或其他一些参与胆固醇合成的中间代谢物浓度的变化。当细胞内的胆固醇或甲羟戊酸等物质的浓度较高的时候,它们能够与上述结构域结合而诱导酶的构象发生变化,从而激活细胞内依赖于泛素的蛋白酶体降解系统。
HMG-CoA还原酶的基因表达需要SREBP-2的转录因子,此转录因子能够对细胞内固醇的浓度做出反应。当细胞内固醇的浓度低的时候,SREBP-2 从内质网膜上的前体蛋白中切割释放出来并移位到细胞核作为转录因子而刺激HMG-CoA还原酶和其他一些参与胆固醇合成的酶基因的表达。
高等动物的脂肪动员是一种受激素控制的过程,胰高血糖素和肾上腺素通过Gs蛋白促进脂肪的水解,而胰岛素则抑制脂肪的水解。脂肪被水解成甘油和FFA,释放出来的FFA进入血流,并与血浆中的清蛋白结合,被转移到其他组织细胞内进一步氧化分解。甘油直接通过循环系统进入肝脏或其他细胞被利用。
脂肪的合成就是甘油的三个羟基被脂酰化的过程。在细胞内,游离的脂肪酸和甘油在缩合成酯之前分别需要被活化成脂酰CoA和甘油-3-磷酸。在脂酰基转移酶催化下,甘油磷酸上两个游离的羟基和脂酰CoA先后缩合形成磷脂酸,后者在磷酸酶的催化下被水解为DAG。在脂酰基转移酶的催化下,DAG上游离的羟基被脂酰化成脂肪。
甘油磷脂的水解由磷脂酶催化。不同的磷脂酶对于甘油磷脂上酯键的特异性不同。鞘磷脂的水解是在溶酶体内进行的,参与水解的酶有鞘脂酶和神经酰胺酶。
甘油磷脂的合成在粗面内质网上进行,其中前二个阶段的反应与脂肪的合成完全一样,而第三阶段的反应有两种方式,都需要消耗CTP。第一种方式首先是磷脂酸被激活成CDP二酰甘油,然后在相应的合酶催化下,与非活化的头部X基团起反应生成各种相应的甘油磷脂同时释放出CMP;第二种方式首先是磷脂酸被水解成1,2-二酰甘油,X基团被活化成CDP-X,然后在相应的合酶的催化下产生甘油磷脂同时释放出CMP。
鞘磷脂的合成同样在粗面内质网上进行,由四步反应组成:二氢鞘氨醇的合成、脂酰基的加入形成N-脂酰基二氢鞘氨醇、N-脂酰基二氢鞘氨醇去饱和成为N-脂酰基鞘氨醇、活化的头部基团结合并最终形成鞘磷脂释放出CMP。
缩醛磷脂的合成场所主要在过氧化物酶体。
鞘糖脂的水解发生在溶酶体,有一系列的酸性水解酶参与,整个反应是高度有序的。鞘糖脂合成的多数反应与鞘磷脂的合成是一样的,只是在神经酰胺形成以后发生一系列的糖基转移反应。在糖基转移反应中,糖基供体通常为UDP-单糖,少数为CMP-单糖。
甘油糖脂的合成则与甘油磷脂合成的多数反应一样,只是在DAG形成以后,在特定的糖基转移酶的催化下,其3号位的羟基接受单糖基而形成。
氨基酸分解代谢可分为氨基代谢和碳骨架的代谢。
氨基的代谢有脱氨基、转氨基和联合脱氨基三种方式,其中联合脱氨基是前两种方式的组合。参与脱氨基反应的主要有L-氨基酸氧化酶、D-氨基酸氧化酶和谷氨酸脱氢酶,其中前两种酶属于黄素蛋白,分别作用于L-氨基酸和D-氨基酸,脱下的氢经过FMN或FAD直接与O2结合形成H2O2。参与氨基酸氧化脱氨基的主要酶是谷氨酸脱氢酶,位于线粒体基质,反应的辅酶既可以是辅酶Ⅰ也可以是辅酶Ⅱ。
7
转氨基反应是指一种氨基酸的氨基被转移到一种α- 酮酸的酮基上形成一种新的氨基酸和一种新的α- 酮酸的过程,由转氨酶催化。转氨反应是完全可逆的,既参与氨基酸的降解,又参与氨基酸的合成。每一种转氨酶都需要磷酸吡哆醛作为辅基,而且绝大多数转氨酶以谷氨酸作为氨基的供体或者以α- 酮戊二酸为氨基的受体。最常见的转氨酶是谷丙转氨酶和谷草转氨酶。Thr、Pro和Lys不能参与转氨反应。
联合脱氨基反应是由转氨酶和谷氨酸脱氢酶组合在一起的脱氨基反应。转氨酶先负责催化将一种氨基酸的氨基转移到α- 酮戊二酸的酮基上,形成谷氨酸,然后谷氨酸脱氢酶催化氧化脱氨基反应,产生α- 酮戊二酸、NH+和NAD(P)H。 4
氨对生物体是有毒的,不同种类的生物体在解除氨的毒害的方式上是不一样的,它与一种生物对水的可得性有关,有的直接排出体外,有的转变为酰胺,有的转变为尿素或尿酸。植物通常使用Asn作为解除氨毒的手段,人类和其他所有胎盘类哺乳动物解除氨毒的方式是将氨转变为尿素,而爬行动物和鸟类则是将氨转变为尿酸。
尿素是通过尿素循环产生的,完整的尿素循环仅存在于肝细胞。尿素循环中的一部分反应发生在肝细胞的细胞液,另一部分反应部分发生在肝细胞的线粒体基质。合成1分子尿素需要消耗4分子ATP,在形成的尿素分子中,1个N原子来自Glu,另一个来自Asp。
碳骨架的代谢有五种方式:严格生糖、严格生酮、生糖兼生酮、经TCA循环彻底氧化成CO2和H2O、重新转变为氨基酸。
生物圈内的氮有N2、NO-、NO-、NH+和氨基酸等几种不同的形式,它们之间是可以相互324
转变的,构成N循环。所有的生物N都以还原形式存在,而主要的无机N以氧化形式存在。生物体获取N需要将氧化形式的N(N2和NO-)转变成还原形式的N(NH+),这几乎都发34
生在微生物和绿色植物中,动物从食物中获取氧化形式的N。
硝酸(盐)的同化由两步反应组成:硝酸(盐)接受2个电子被还原成亚硝酸(盐)和亚硝酸(盐)接受6个电子被还原成NH+。N2固定仅发生在某些原核生物。所有的生物固氮系4
统都由固氮酶复合物催化,固氮酶复合物由固氮酶还原酶和固氮酶组成。整个固氮反应中,每1对电子传递需要4个ATP,每固定1分子N2需要消耗16个ATP。
不同生物体合成氨基酸的能力是不一样的,植物和微生物在有合适的N源时,能够从头合成所有的20 种常见氨基酸,而哺乳动物只能制造其中的10种。任何氨基酸的合成的前体都来自于糖酵解、TCA循环或磷酸戊糖途径,其中N原子是通过Glu或Gln进入相关的合成途径的,而合成的场所有的在细胞液,有的在线粒体。
按照各氨基酸合成前体的性质,所有的氨基酸可分为5大家族:① α- 酮戊二酸家族这一族氨基酸以α- 酮戊二酸作为合成的前体,包括Glu、Gln、Pro和Arg,在某些生物体还包括 Lys;② 天冬氨酸家族属于这一族的氨基酸有Asp,Asn,Lys,Met,Thr和Ile;③ 丙酮酸家族属于这一族的氨基酸有Ala,Val,和Leu;④ 甘油酸-3-磷酸家族属于这一族的氨基酸有Ser、Gly和Cys;⑤ 芳香族氨基酸Phe、Tyr,和Trp的合成是细胞通过从头合成和补救合成两种途径合成核苷酸。从头合成从最简单的小分子开始,经过多步反应,消耗更多的能量最后生成核苷酸;补救途径是指核苷酸降解的中间产物包括核苷和嘌呤重新被利用并转变为核苷酸的过程,涉及的反应和能耗较少。
嘌呤核苷酸从头合成的小分子前体包括CO2、Asp、Gly、Gln和N 10-甲酰四氢叶酸,反应中消耗ATP或GTP。整个合成途径由10多步反应构成。PRPP的合成是第一步反应。PRPP不仅参与嘌呤核苷酸和嘧啶核苷酸的从头合成,还参与它们的补救合成途径。5-磷酸-1-β-核糖胺的形成是第二步反应,由谷氨酰胺:PRPP氨基转移酶催化。该反应是 8
嘌呤核苷酸从头合成的限速步骤,受高度调控,调节的方式是有活性的单体和无活性的二聚体的互变。IMP、GMP和AMP促进单体向二聚体的转变而反馈抑制此步反应,反应的底物PRPP则激活该酶的活性。余下的反应从磷酸核糖胺直到IMP共由9步反应组成。然后再由IMP分别转变成AMP和GMP。在IMP转变为AMP的反应中,消耗GTP,而在IMP转变为GMP的反应中,消耗的是ATP,这样可以有效地调节细胞内GTP和ATP之间的比例。
嘧啶核苷酸从头合成的前体包括Gln、CO2和Asp等,是先得到UMP,然后再在UTP的水平上得到CTP。嘧啶核苷酸的从头合成是先形成嘧啶环,然后再与PRPP形成β-N-糖苷键,其中有些反应是在线粒体内发生的。氨甲酰磷酸的合成是第一步反应,由CPS-Ⅱ催化,第二步反应由天冬氨酸转氨甲酰基酶催化。细菌嘧啶核苷酸合成的限速酶为天冬氨酸转氨甲酰基酶,其中CTP和UTP为它的反馈抑制剂,ATP为别构激活剂;哺乳动物嘧啶核苷酸合成的限速酶是CPS-Ⅱ,UDP或UTP抑制,PRPP激活。EGF能够诱导CPS-Ⅱ的磷酸化,使其降低对UTP抑制的敏感性,但增强了对PRPP激活的敏感性。
参与嘌呤核苷酸的补救合成的酶有两类:一类属于嘌呤碱基腺嘌呤磷酸核糖转移酶,需要嘌呤碱基和PRPP,它包括APRT和HGPRT,前者用于AMP的补救合成,后者用于IMP和GMP的补救合成;另一类包括核苷磷酸化酶和核苷激酶。参与嘧啶核苷酸补救合成的酶主要有核苷磷酸化酶和核苷激酶。许多核苷类药物都是通过此途径转变为相应的核苷酸的。
脱氧核苷酸是在核苷酸还原酶的催化下由核糖核苷酸还原而成。最普遍是NDP还原酶,反应机制涉及活性中心的一个Tyr残基自由基,还需要一个双铁氧桥的帮助,以维持上述自由基的再生。
大肠杆菌使用NDP还原酶,直接的还原剂为硫氧还蛋白,而最终的还原剂为NADPH。其中NADPH在硫氧还蛋白还原酶的催化下负责还原性硫氧还蛋白的再生。NDP还原酶活性受到严格的调控。大肠杆菌的NDP还原酶除了活性中心以外,还有S位点和A位点。这两个别构位点与一系列别构效应物的相互作用构成了极为精妙的调节网络,从而维持了细胞内脱氧核苷酸和核糖核苷酸之间以及四种脱氧核苷酸之间的平衡。
胸苷酸是在胸苷酸合酶的催化下,由dUMP甲基化而合成的,甲基供体为N5,N10-亚甲基四氢叶酸。dUMP由dUTP水解而成。
在核苷酸酶、核苷酶或核苷磷酸化酶依次催化下,核苷酸可分解成碱基和核糖,碱基还可以进一步分解,核糖则可融入糖代谢。嘌呤核苷酸在某些动物体内分解的终产物是尿酸,在其他生物,尿酸还可能进一步分解成尿囊素或尿囊酸或乙醛酸和尿素。
痛风是尿酸过量产生或者尿酸排泄不畅引起。治疗痛风的特效药物是别嘌呤醇,它是黄嘌呤氧化酶的一种自杀性底物。SCID是ADA基因有缺陷引起的,可使用骨髓移植治疗或基因治疗。Lesch-Nyhan综合征是HGPRT有缺陷造成的。乳清酸尿症的病因是具有乳清酸磷酸核糖转移酶和OMP脱羧酶活性的双功能酶的缺陷。
抗核酸代谢类药物通常作为竞争性抑制剂或自杀性底物来抑制核苷酸合成的某一种酶,干扰或阻断核苷酸的合成,从而阻断DNA的复制和转录,致使细胞的分裂受到抑制。抗核酸代谢类药物除了可用于治疗癌症以外,还经常用作抗病毒的药物。一般的抗核酸代谢类药物可分为叶酸类似物、谷氨酰胺类似物、碱基类似物、核苷类似物以及核苷酸类似物。
通过莽草酸途径。His的合成与Trp的合成相似,与核苷酸合成途径一样需要PRPP。
9