广州大学学生动物学实验报告
开课学院及实验室:生科院生化楼603 20##年9月10日

第二篇:动物生理学课件
动物生理学
理论讲稿
绪言
第一章 机体的基本生理过程
第二章 血液生理
第三章 神经生理
第四章 血液循环生理
第五章 呼吸生理
第六章 消化生理
第七章 能量代谢生理
第八章 泌尿生理
第九章 内分泌生理
第十章 生殖生理
第十一章 泌乳生理
生理学的发展
绪言
- 家畜生理学的研究对象、任务、方法和分类
一、对象
- 家畜生理学:家畜生理学是生理学的一个分支,它是研究健康家畜(包括家禽)所表现的正常生命现象或生理活动及其规律性的科学。
在农业范畴内,家畜生理学研究对象主要包括畜、禽和鱼类,除了分别阐明哺乳动物(家畜)、鸟类(家禽)及脊椎动物(鱼类)的共性的生理学外,还应该研究与其生产性能紧密联系的生理学特性。例如,畜禽经长期驯养选育,生产性能显著提高,乳牛一个泌乳期一般可产乳6000-7000公斤,蛋用鸡年产量300枚以上,均超过其祖先或原始品种数十倍。因此,机体生理上必然发生变化,出现若干特殊规律性。
家畜是由各器官系统构成的完整统一体,机体内的个别器官系统的生理活动,都与其它生理活动保持着复杂而精确的联系和相互制约的关系,也都适应于家畜当时的整体生理活动的情况,另一方面,机体各器官系统得生理活动,又是以构成这些器官系统的细胞及其内部的物质分子的生物学特性为基础的。
- 生理学研究的三种不同水平
- 分子和细胞生理学:即研究细胞及其物质分子结构与功能的关系,以及它们内部所发生的各种生物化学变化和生物物理变化的过程和规律。
- 器官和系统生理学:即研究各器官系统所表现的各种特殊生理活动的过程和规律,以及它们之间的相互关系。
- 整体和环境生理学:即从家畜的整体观点出发,研究机体对全部生理活动的调节、整合过程和规律、以及机体与生活环境的辩证统一关系。
二、基本任务
(1)阐明细胞、组织、器官及整个机体水平上生命活动的规律性。
(2)研究机体与周围环境相互作用机理。
(3)动物在进化和个体发育阶段的机能。
(4)各种动物生理学质的差别。
(5)动物生理学还由于研究特殊的物质运动-----生命,为科学的辩证主义提供科学依据。
三、研究方法
1、慢性实验:是在无菌条件下对健康动物进行手术,暴露要研究的器官(如消化道各种造瘘手术)或摘除、破坏某一器官(如切除某一内分泌腺),然后在接近正常生活的条件下,观察所暴露器官的某些功能,观察摘除或破坏某器官后所产生的功能紊乱等。
2、急性实验
(1)在体(in vivo) 实验:是在无痛条件下剖开动物身体,对某一两个器官进行实验观察。
(2)离体(in vitro) 实验:是在动物体内取出某一器官,置于人工的环境中,研究它的机能和有关因素的作用。
第一章机体的基本生理过程
第一节稳态与机体机能活动的调节
一、生命的基本特征
1.新陈代谢(metabolism)新陈代谢是宇宙间普遍的永远不可抵抗的规律,它是宇宙间一切事物运动发展的共同的本质,生物有机体也不可能违背这一规律。生物体生活在适宜环境中,总是在不断的重新建造自身的特殊结构,虽然从生物体的外表可能看不出什么显著的变化,但实际上,它的各个部分都在不断地合成新分子代替旧的,这个过程就称新陈代谢。即机体主动地与环境进行物质和能量交换,以及机体内部物质和能量的转变转移过程。
2.兴奋性(excitability)一切活组织或细胞,当其周围环境条件迅速改变时,有产生动作电位并发生反应(response)的能力或特性,称为兴奋性或应激性(感应性,irritability)。这种引起反应的环境条件的迅速变化称为刺激(stimulus)。
在动物组织中,通常以神经细胞和肌细胞以及腺细胞表现出较高的兴奋性,因此被称为可兴奋细胞或可兴奋组织(excitable tissue)。
活组织在接受刺激而发生反应时,其表现形式有两种:一种是由相对静止状态转变为显著活动状态,或由活动弱变为活动强,称为兴奋(excitation);另一种是由显著活动状态转变为相对静止状态,或由活动强变为活动弱,称为抑制(inhibition)。
3.生殖(reproduction)
生殖(自我复制,self-replication)是个体生长发育到一定阶段时可产生相似的另一新个体的过程。
二、内环境与稳态
1.内环境(internal environment)高等动物特别是哺乳动物大都生存于外界大气环境之中,但是有机体的大多数细胞并不直接与外界环境发生接触,也不能与外环境直接进行物质交换。有机体的大多数细胞实际上是浸浴在包括血浆、淋巴液、组织液等组成的细胞外液(extracellular fluid)中,因而称细胞外液为机体的内环境。
2.稳态(homeostasis)及其生理意义有机体通过内环境与外环境进行物质交换并不断地代谢,因此内环境的成分和理化性质不是固定不变的,而是在一定范围内波动,保持着动态的平衡。这种动态平衡状态称为稳态,又叫自稳态。
稳态的维持具有重要的生理意义:
①稳态是新陈代谢的必要保证,如果细胞外液的温度或pH值等发生变化将改变有关酶的活性,从而影响体内各种酶促反应过程;又如水溶液渗透压的变化将影响血管内外、细胞内外的水平衡,也将影响物质交换过程。
②细胞正常兴奋性的维持需要膜内外离子浓度的相对稳定。
③在外界环境剧烈变化(如温度)时,内环境保持相对稳定是机体具有适应能力的前提。
3.生物节律(biorhythm) 从生理指标到整体机能都不是固定不变,也不是匀速运动,往往表现出周期性变化的规律。在生理学中,把按一定的时间顺序发生变化,即按一定的时间重复出现、周而复始的功能活动变化节律叫做生物节律。
三、机体机能活动的调节
(一)机体机能活动的调节方式
1.神经调节(neuroregulation)
神经调节是指机体在刺激的作用下,通过神经系统的反射活动调节器官的机能。神经调节的基本方式是反射。

 (1)、反射(reflex):在中枢神经系统参与下,机体对内、外环境刺激所发生的反应,这种调节过程称反射。如蚊子叮咬打掉,事物入口唾液分泌、消化液分泌。
(1)、反射(reflex):在中枢神经系统参与下,机体对内、外环境刺激所发生的反应,这种调节过程称反射。如蚊子叮咬打掉,事物入口唾液分泌、消化液分泌。
(2)、反射弧(reflex arc):实现反射的结构单位和功能单位。包括感受器、传入神经、神经中枢、传出神经和效应器五个部分。
(3)、感受器(Receptor):是接受刺激的装置,任何一个反射活动首先从它开始,能把体内、外各种刺激感受到,然后把刺激转变为神经冲动。
(4)、传入神经(Afferent nerve):连接感受器和反射中枢的N质。
(5)、神经中枢(Central neurous):指脑或脊髓内一定部位执行某种机能的NC群,能对传入的信号进行分析综合。简单反射的中枢位于低级部位(脊髓、延髓,如排尿、排粪),复杂的反射中枢位于NS的高级部位(大脑皮质,如思维)。
(6)、传出神经(Efferent nerve):是将感受器和效应器与神经中枢联系起来的通路。
效应器(Effector):是产生反应的结构;根据中枢指令发生相适应反应的器官,环节破坏,反射不出现,紊乱。
特点:迅速而精确,作用部位准确,持续时间较短。
2.体液调节(humoral regulation)
机体的内分泌细胞能产生某些特异性化学物质(如激素),通过血液循环输送到全身各处,对某些特定的组织起作用,以调节机体的新陈代谢、生长、发育、生殖等机能活动,这种激素调节就属于体液调节。
特点:出现缓慢,作用部位较广泛,持续时间较长。
在人和大多数高等动物具有神经调节和体液调节两种机制,二者相辅相成,共同完成机体机能调节的任务。大多数内分泌腺也直接或间接地受到神经系统的控制,从而使体液调节成为神经调节的一环,相当于反射弧传出路径上的一个延续部分,这种情况称为神经—体液调节(neurohumoral regulation)。因此,可以认为神经调节是高等动物机体最主要的调节方式,处于主导地位。
3.自身调节(autoregulation)
是指细胞、组织、器官在不依赖于外来的神经或体液调节情况下,自身对刺激发生的适应性反应过程。如回心血量增加时,心肌细胞初长度增大,心肌收缩力量增强,使心脏中血容量变化不大。一般说来,自身调节的调节幅度较小,也不十分灵敏,但对生理功能的调节仍有一定意义。
(二)机体机能活动的调控模式
1.非自动控制系统象各种有害刺激引起动物应激这样的调节活动,刺激决定着反应,而反应不能改变控制部分的活动,属开环系统。这种控制系统无自动控制的能力,只要刺激存在,反应就将继续下去。非自动控制系统在体内不多见。
2.反馈(feedback)控制系统由控制部分(中枢)发出的信息可改变受控部分(效应器)的状态,而受控部分反过来又发出信息、把接受控制的状态结果不断地报告给控制中抠,使控制中枢得以参照实际情况不断纠正和调整发出的信息,以达到对受控部分精确的调节,是闭环系统。这种由受控部分送回到控制中枢的信息称为“反馈”信息,这种调节方式称为反馈调节。
负反馈(negative feedback):如果反馈信息可使控制中枢的初始控制信息减弱,称为负反馈。
正反馈(positive feedback):如果可使控制中枢的原始信息加强,则称为正反馈。
3.前馈(feedforward)控制系统所有条件反射均属于前馈控制系统。在前馈控制系统中监测装置(感受器)不是检测输出变量的波动,而是直接检测到干扰信息(条件刺激)后发出前馈信息,作用于控制系统,调整控制信息以对抗干扰信息对受控系统的作用,从而使输出变量保持稳定。它可以避免反馈调节的波动性和反应滞后,更好地保持稳态,其预见性更具有适应性意义。但前馈控制引起的反应也有可能失误,如见到而未吃到食物时的唾液分泌。
第二节细胞的基本功能
一、细胞膜结构及物质转运功能
细胞膜是细胞表面一层连续而封闭的界膜,又称质膜或浆膜,细胞内部也存在着类似细胞膜的膜性结构,围成各种细胞器,实现细胞内空间上的区域化和功能上的有序化。因此,膜的三层结构被认为是细胞中普遍存在的基本结构,统称为单位膜或生物膜。
(一)膜的化学组成和分子结构
在电子显微镜下分为三层的各种膜性结构主要由脂类、蛋白质和糖类等物质组成,一般都是以蛋白质和脂质为主,糖类只占极少量。
目前流体镶嵌模型(fluid mosaic model)得到较多实验事实支持并为大多数人所接受。这一假想模型的基本内容是:膜的共同结构特点是以液态的脂质双分子层为基架,其中镶嵌着具有不同分子结构和不同生理功能的蛋白质,后者主要以α-螺旋或球形蛋白质的形式存在。
1.脂质双分子层(lipid bilayar)膜的脂质中以磷脂类为主,约占脂质总量的70%以上;其次是胆固醇,一般低于30%;还有少量属鞘脂类的脂质。近年来发现,膜结构中含量相当少的磷脂酰肌醇,几乎全部分布在膜的靠胞浆侧;这种脂质与细胞接受外界影响、并把信息传递到细胞内的过程有关。
磷脂的基本结构是:一分子甘油的两个羟基同两分子脂酸相结合,另一个羟基则同一分子磷酸结合,后者再同一个碱基结合。根据这个碱基的不同,动物细胞膜中的磷脂主要有四种(图1-2):磷脂酰胆碱、磷脂酰乙醇胺、磷脂酰丝氨酸和磷脂酰肌醇。所有的膜脂质都是一些双嗜性分子,磷脂的一端的磷酸和碱基是亲水性极性基团,另一端的长烃链则属疏水性非极性基团。鞘脂类的基本结构和磷脂类似,但不含甘油。胆固醇结构很特殊,它含有一个甾体结构(环戊烷多氢菲)和一个8碳支链。通过对红细胞膜所作的化学测定和计算,提出的双分子层模型中,每个磷脂分子中的极性基团都朝向膜的外表面或内表面,而非极性基团则在膜的内部两两相对。脂质分子的这种定向而整齐的排列,从热力学角度分析,包含的自由能最低,因而最为稳定,可以自动形成和维持。根据同样的原理,如果让脂质分子在水溶液中受到激烈扰动时,脂质有可能形成含水的小囊,但这囊只能是由脂质双分子层形成。这种人工形成的膜囊,称为脂质小体,似人造的细胞空壳,有很大的理论研究和实用价值。
脂质的熔点较低,膜中脂质分子在一般体温条件下是呈液态的,即膜具有某种程度的流动性。脂质双分子层在热力学上的稳定性和它的流动性,能够说明何以细胞可以承受相当大的张力和外形改变而不致破裂,而且即使膜结构有时发生一些较小的断裂,也可以自动融合而修复,仍保持连续的双分子层的形式。当然,膜的这些特性还同膜中蛋白质和膜内侧某些特殊结构(称为细胞骨架)的作用有关。应该指出的是,膜的流动性一般只允许脂质分子在同一分子层内作横向运动,由于分子的双嗜性,要脂质分子在同一分子层内作“掉头”运动,或由一侧脂质层移到另一侧脂质层,这意味着有极性的磷酸和碱基的一端要穿越膜内部的疏水性部分,这是不容易或要耗能的。
2.膜蛋白质 膜蛋白质主要以两种形式同膜脂质相结合:有些蛋白质以其肽链中带电的氨基酸或基团,与膜两侧的脂质极性基团相互吸引,使蛋白质分子像是附着在膜的表面,这称为表面蛋白质;有些蛋白质分子的肽链则可以一次或反复多次贯穿整个脂质双分子层,疏水的α-螺旋正好与膜内疏水性烃基相吸引,肽链两端露出在膜的两侧,这称为结合蛋白质。膜中的蛋白质,具有不同的分子结构和功能。生物膜所具有的各种功能,在很大的程度上决定于膜所含的蛋白质;细胞和周围环境之间的物质、能量和信息交换,大都与细胞膜上的蛋白质分子有关。
由于脂质双分子层是液态的,镶嵌在脂质层中的蛋白质是可移动的,即蛋白质分子可以在膜脂质分子间横向漂浮移位;不同细胞膜中的不同蛋白质分子的移动和所在位置,存在着精细的调控机制。例如,骨骼肌细胞膜中与神经肌肉间信息传递有关的通道蛋白质分子,通常都集中在肌细胞膜与神经末梢分布相对应的那些部分(终板膜);而在肾小管和消化管上皮细胞,与管腔相对的膜和其余部分的膜中所含的蛋白质种类大不相同,说明各种功能蛋白质分子并不都能在所在的细胞膜中自由移动和随机分布,而实际存在着有区域特性的分布,显然同蛋白质完成其特殊功能有关。膜内侧的细胞骨架可能对某种蛋白质分子局限在膜的某一特殊部分起着重要作用。
3.细胞膜糖类细胞膜所含糖类甚少,主要是一些寡糖和多糖链,它们都以共价键的形式和膜脂质或蛋白质结合,形成糖脂和糖蛋白;这些糖链绝大多数是裸露在膜的外面一侧的。这些糖链的意义之一在于以其单糖排列顺序上的特异性,作为它们所在细胞或它们所结合的蛋白质的特异性“标志”。例如,有些糖链可以作为抗原决定簇,表示某种免疫信息;有些是作为膜受体的“可识别”部分,能特异地与某种递质、激素或其他化学信号分子相结合。如人的红细胞ABO血型系统中,红细胞的不同抗原特性就是由结合在膜脂质的鞘氨醇分子上的寡糖链所决定的,A型抗原和B型抗原的差别仅在于此糖链中一个糖基的不同。由此可见,生物体内不仅是多聚核苷酸中的碱基排列顺序和肽链中氨基酸的排列顺序可以起“分子语言”的作用,而且有些糖类物质中所含糖基序列的不同也可起类似的作用。
(二)细胞膜的跨膜物质转运功能
细胞膜具有选择通透性(permeability),即不同物质通过细胞膜的难易程度有很大区别。因此又把生物膜叫半透膜。根据跨膜物质转运过程中物质运动的方向和是否需供给能量,基本上可分为被动转运(passive transport)和主动转运(active transport)两大类)。
1.被动转运物质从高浓度的一侧,通过细胞膜运送到低浓度的一侧,即顺浓度的方向跨膜转运的过程称为被动转运。该过程消耗了高浓度溶液的化学势能,是一种不需要细胞提供能量的自发过程。
特点:物质的转运速率既依赖于膜两侧被转运物的浓度差,又与被转运物质的分子大小、电荷和在脂质层中的溶解性有关。被动转运包括单纯扩散(simple diffusion)和易化扩散(facilitated diffusion)。
⑴单纯扩散在生物体系中,只有O2、N2、苯等疏水性小分子及CO2、甘油、尿素等小的极性分子能迅速地经扩散通过脂双层膜。这种可以认为是单纯的物理过程的跨膜物质转运方式称为单纯扩散。
⑵易化扩散稍大些的极性分子(如葡萄糖、氨基酸)和小的带电离子通过细胞膜需要借助膜蛋白的帮助,扩散速率显著增大,称为易化扩散。
2.主动转运细胞之所以能维持恒定的离子梯度差,是由于细胞膜具有逆浓度梯度转运物质的功能,称为主动转运,需要细胞膜或膜所在细胞供给能量。Na+ 被运到膜外、K+ 被运进细胞就是由一种被称为钠钾泵(又简称钠泵,sodium pump)的膜蛋白完成的主动转运。
钠泵由催化亚单位和一个糖蛋白以四聚体形式存在于膜上。钠泵的活动需要Na+和K+的特定分布(Na+在膜内、K+位于膜外)时才能分解胞内的ATP提供能量而进行,因此又被称为Na+-K+依赖式ATP酶。
钠泵的作用机理可:①钠泵与膜内Na+结合,②酶被激活并在细胞质侧水解ATP,泵本身被磷酸化,③其诱导的构象变化运送Na+通过膜并在外侧释放,④此时的泵与膜外K+结合,⑤泵去磷酸化,⑥钠泵回到原来的构象,并运送K+通过膜释放到胞内。
钠泵活动具有重要的生理意义,首先细胞内高K+水平是许多代谢反应的必备条件;其次维持细胞内外离子不均匀分布,建立起一种势能贮备可以完成许多生理功能。有研究表明,静止动物消耗ATP的1/3用于离子泵的活动。可以说主动转运是机体最重要的物质转运形式。除钠钾泵外,体内类似的还有钙泵(Ca2+-Mg2+依赖式ATP酶)、质子泵(H+-K+依赖式ATP酶)等,分别与肌细胞收缩和胃酸分泌等生理活动有关。
3.胞吐(exocytosis)与入胞(endocytosis)式物质转运以上所述的是小分子或离子的跨膜转运方式,对于生物大分子和固态或液态的物质团块来说,则是通过更加复杂的结构和功能的改变实现的。如神经递质的释放和许多细胞分泌蛋白质等,细胞内物质先被囊泡裹入形成分泌泡,然后与细胞膜接触、融合并向外释放被裹入的物质,这个过程称为胞吐、出胞或外排作用。在分泌泡与质膜融合这一关键步骤中,由胞外进来的Ca2+起到了重要作用。
细胞从外界吸附的大分子或颗粒,逐渐被细胞膜的一小部分包围、内陷,其后从细胞膜上脱落下来而形成含有摄入物质的细胞内囊泡的过程,称为入胞或内吞作用。
入胞作用又可分为以下3种形式:
吞噬作用(phagocytosis):胞摄取的是固体物质,可形成大的囊泡。
胞饮作用(pinocytosis):小的囊泡形式将细胞周围的微滴状液体吞入细胞内的过程。
受体介导式入胞(receptor-mediated endocytosis):内吞物(配体)与细胞表面的专一性膜蛋白(受体)结合为起始的入胞。
二、细胞的兴奋性与生物电现象
(一)细胞生物电现象
生命活动过程中出现的电现象称为生物电现象(bioelectric phonomenon)。
1.细胞的静息电位(resting potential)和动作电位(action potential)
⑴静息电位细胞未受刺激时存在于细胞膜内外两侧的电位差,称为静息电位或膜电位(membrane potential)。通常呈膜外为正、膜内为负的极化(polarization)状态,如动物体内神经细胞和肌细胞有较大的静息电位,约-65mV至-100mV之间;相对最小的是红细胞,仅为-10mV。
⑵动作电位可兴奋组织受到刺激而发生兴奋时,细胞膜原来的极化状态迅速消失,并继而发生倒转和复原等一系列电位变化,称为动作电位。
神经细胞的动作电位可大体看作两部分:开始是一个短促而尖锐的脉冲,习惯称锋电位(spike potential);其后是在静息电位水平上下的微小而缓慢的电位波动—后电位(after-potential)。锋电位包括了从静息电位到零电位(局部膜瞬间内外电位相等)的去极化、从零电位再向上的反极化(也称超射,overshoot)和恢复极化的下降支3个阶段。后电位包括负后电位和正后电位(超极化状态)两个阶段。锋电位尤其是其上升支表示细胞处于兴奋状态,通常所说的神经冲动(impulse)就是指一个个沿着神经纤维传导的动作电位或锋电位。
2.生物电现象产生的机理膜内外两侧离子分布的不均匀及细胞膜的选择通透性是生物电现象产生的基础。
⑴静息电位产生的机制静息状态下,细胞膜主要对K+有通透性,在浓度差作用下K+外流。带正电荷的K+外流吸引了带负电的蛋白质分子有外流倾向。但膜对蛋白质不通透,蛋白质被阻隔积累在膜的内表面,相应地吸引流出的K+集中分布于膜的外表面,这样产生外正内负的极化状态。因此说细胞静息电位基本上相当于K+外流产生的跨膜平衡电位。
⑵动作电位产生的机制是膜在受到刺激时Na+通透性的突然增大,以致超过K+通透性,使大量Na+涌入膜内的结果。
现在以神经细胞为例说明动作电位产生的完整过程。外来刺激先引起膜对Na+通透性突然增大,Na+大量内流,导致膜电位迅速去极化乃至反极化,构成动作电位的上升支,时间少于1ms;在此过程中Na+通道很快关闭而K+通道(不同于形成静息电位时的K+泄漏通道)大量开放,膜内K+大量外流使膜电位迅速恢复,构成动作电位下降支(图1-9 电导反映膜的通透性)。负后电位表示迅速涌出的K+蓄积在膜外侧附近,暂时阻碍了K+进一步外流所致复极化变慢,而正后电位是由于膜上钠钾泵启动,在恢复膜两侧离子分布状态过程中生电性作用(泵出3个Na+同时泵入2个K+,造成超极化)等原因引起,与兴奋后的恢复有关。
(二)兴奋性和刺激引起兴奋的条件
1.刺激引起兴奋的条件自然界中能引起机体反应的刺激是多种多样的,按其性质可分为物理性刺激如声、光、电、机械、温度等,化学性刺激如酸、碱、各种化学物质等和生物刺激如细菌、病毒等。对机体某种感受器来说,刺激有适宜与不适宜的区别,如声波是听觉器官、光波是视觉器官的适宜刺激(optimal stimulus)等。应该指出,这种分工是长期进化过程中器官功能特殊化的结果,但并不是某种器官只有唯一的适宜刺激。所谓适宜刺激是指感受器对该性质的变化最敏感(即阈值最低)、或引起兴奋所需能量最低。在诸多性质的刺激中,电变化具有可定量调节、易重复、对组织损伤小等优点,为生理实验所常用。
即使是适宜刺激,若在强度、作用时间和强度—时间变化率三个要素的某一方面达不到某一临界值,还是不能引起兴奋。如用干电池电流刺激肌肉,通电和断电的瞬间可引起收缩;接通状态下则无反应,因为此时的强度—时间变化率为零之故。这三个参数并不是固定值,如果其中一个或两个参数变了,其余的也会发生相应改变。为了研究各参数间的相互关系,可先固定其中某一个参数,然后观察另外两个参数的相互影响。例如,使用方波电脉冲刺激“神经—肌肉标本”以肌肉收缩作为神经兴奋的间接指标时,由于不同大小和持续时间的方波其斜率都是一样的(方波的振幅代表强度、波宽代表时间),因而认为这类刺激的强度—时间变化率固定不变,就可分析刺激强度和作用时间的相互影响和关系。实验结果表明,在强度—时间变化率固定不变的情况下,在一定范围内,引起组织兴奋所需的最小刺激强度与该刺激的作用时间呈反变关系。就是说,当所用的刺激强度较大时,引起组织兴奋只需用较短的作用时间;而当刺激强度较小时,需用较长的作用时间才能引起组织产生兴奋。如果把能够引起兴奋的不同刺激强度和相对应的作用时间描绘在坐标纸上,便可得到一条近似于双曲线的曲线,称为强度—时间曲线(strength-duration curve)。强度—时间曲线上的任何一点都表示一个刚能引起组织兴奋的最小刺激,称为阈刺激(threshold stimulus),曲线右上方各点表示阈上刺激,左下方各点表示阈下刺激。不同的组织描绘出的强度—时间曲线不同,如同为神经纤维,较细的神经纤维测得的强度—时间曲线偏右上方(图1-11中虚线),说明兴奋性较低。强度—时间曲线能够较全面地反映组织细胞的兴奋性,但是当兴奋性发生迅速变化时,要测得一条强度—时间曲线实际上有困难。因此有人主张用时值(chronaxie法文)代替。如图1-11所示,时值正位于强度-时间曲线中部曲度最明显的部位,能较好地反映整个曲线的位置,是衡量组织兴奋性高低的又一指标。但测定时值也非易事,实际工作中衡量组织兴奋性常用而简便的指标是阈强度(threshold intensity)。即固定刺激的强度—时间变化率和作用时间,测定引起组织兴奋所需的最小刺激强度,又简称阈值(threshold)。引起某组织兴奋所需的阈强度愈小,说明该组织兴奋性愈高;反之,阈强度愈大,说明组织兴奋性愈低。
2.组织兴奋性的变化
以神经组织为例,兴奋后首先出现一个非常短暂的绝对不应期(absolute refractory period),然后出现相对不应期(relative refractory period)。继而是超常期(supranormal period)和低常期(subnormal period)。
3.阈电位(threshold potential)和锋电位的引起外来刺激引起兴奋,即产生动作电位的过程已比较清楚。如神经细胞受到电刺激时,先引起局部膜的去极化,膜对Na+通透性增大,Na+内流。当外来刺激较小时,Na+内流的去极化倾向可被K+外流所纠正,只会发生局部微小的电位波动;只有外来刺激足够大,能使静息电位的绝对值减小到一个临界值(称为阈电位)时,Na+通道大量开放,Na+大量内流,膜电位进一步去极化。如此反复促进形成一种正反馈过程,称为Na+内流的再生性循环(或Hodgkin循环,图1-13),其结果去极化迅速,形成锋电位陡峭的上升支,直至Na+平衡电位。由此可见,阈电位不是单一通道的属性,而是在一段膜上能使通道开放的数目足以引起上述再生性循环出现的膜内去极化的临界水平。因此,只要外来刺激大于能引起再生性循环的水平,膜内去极化的速度就不再决定于原刺激的大小,而取决于原来静息电位的值和膜两侧浓度差。就是说动作电位要么不产生(无),一旦产生就达到最大值(全),符合“全或无定律”(all-or-none law)。
4.局部反应(local response)及其特性细胞受到单个阈下刺激的作用不会产生动作电位,但不是无反应,也将引起去极化,但未达到阈电位水平,仅能使Na+少量通道开放,这时的Na+内流叫做局部反应或局部兴奋。局部兴奋有以下几个基本特性:①不是“全或无”的,而是随着阈下刺激的增大而增大;②只能呈递减性传播,传播数十至数百微米即消失,称为电紧张扩布,这与动作电位截然不同;③由于没有不应期所以具有总和(summation)效应,即多个阈下刺激引起的多个局部反应如果在时间上(在同一部位连续给予)或空间上(同时在相邻的部位给予)叠加起来,就有可能使膜的去极化达到阈电位,从而引发动作电位。体内某些感受器细胞、部分腺细胞和平滑肌细胞,以及神经细胞体上的突触后膜和骨骼肌细胞的终板膜,受到刺激时不是产生动作电位,而只出现原有静息电位的微弱而缓慢的变动,分别称为感受器电位、慢电位、突触后电位和终板电位。它们也具有类似局部兴奋的特性。
(三)兴奋在同一细胞上的传导
动作电位还有一个显著特点是不衰减性传导(conduction)。动作电位一旦在细胞膜的某一部位产生,它就会向整个细胞膜传播,而且它的幅度不会因为传播距离的增加而减弱。例如,一条无髓神经纤维某点发生动作电位,膜出现反极化即外负内正的状态;而相邻部位仍处于内负外正的静息状态。无论细胞外液还是细胞内液都具有良好的导电性,于是已兴奋的神经段与它相邻的未兴奋的神经段之间由于存在电位差而有电荷移动,其方向是膜外的正电荷由未兴奋段移向已兴奋段;膜内的正电荷由已兴奋段移向未兴奋段。相邻段细胞膜去极化并达到阈电位水平,从而爆发动作电位。就这样兴奋膜与相邻未兴奋膜之间产生的局部电流(图1-14A)不断地流动下去,就表现为兴奋(动作电位)在整个细胞上的传导。以上所谓动作电位的传导,实际是已兴奋的膜通过局部电流“刺激”了未兴奋的膜部分,使之产生动作电位。由于锋电位产生期间电位变化的幅度和陡峭度相当大,局部电流的强度超过了引起邻近膜兴奋所需的阈强度数倍以上,因而传导可以相当“安全”而无“阻滞”地继续下去。无髓神经纤维、骨骼肌细胞等基本上都是遵循这样的机制。
有髓神经纤维外面包裹着一层既不导电、离子又不能通过的髓鞘,动作电位只能在没有髓鞘的朗飞结处才能传导,因此动作电位是越过每一段带髓鞘的神经纤维呈跳跃式传导(saltatory conduction,图1-14B),加之有髓神经纤维较粗,电阻较小,所以动作电位在有髓神经纤维上的传导速度要比在无髓神经纤维上快得多。虽然跳跃式传导本质上也是局部电流,但速度快得多。脊椎动物用跳跃传导的方式解决了高速传导神经冲动的问题,而不需要发展粗笨的巨大神经干。
需要指出的是,兴奋的传导是电传导(速度约100~2m/s),不是电子移动的电流传导(速度为3×108m/s),也不是太慢的化学扩散。
三、兴奋的细胞间传递
多细胞间信息传递(transmission)的主要形式是化学性信号。有内分泌或远距分泌、突触传递、神经内分泌、旁分泌和自身分泌等几种类型。
根据已有的研究,跨膜信号传递的方式可概括为由具有特异感受结构的通道蛋白质、受体—G-蛋白—第二信使系统、酪氨酸激酶受体等3种方式。
(一)由具有特异感受结构的通道蛋白质完成的跨膜物质转运
1.化学门控通道在神经-肌肉接头处,当神经末梢有冲动到达时可释放神经递质(neurotransmitter)—乙酰胆碱(ACh),ACh扩散过间隙到运动终板膜,与终板膜上N-ACh化学门控通道(图1-5)的α-亚单位结合,打开通道使Na+和K+跨膜易化扩散,产生终板电位而完成跨膜信号传递。目前已经证明一些氨基酸类神经递质,包括谷氨酸、门冬氨酸、γ-氨基丁酸和甘氨酸等,主要通过类似机制影响其靶细胞。
2.电压门控通道在动物界,除了一些特殊的鱼类,一般不存在专门感受外界电刺激或电场改变的器官或感受细胞,但在体内有很多细胞,如神经细胞和各种肌细胞,在它们的细胞膜中却具有多种电压门控通道蛋白质。它们可由于同一细胞相邻的膜两侧出现的电位改变而出现通道的开放,并由于随之出现的跨膜离子流,而出现这些通道所在膜的特有的跨膜电位改变或细胞内功能变化。如Na+、K+和Ca2+通道即属此类。
3.机械门控通道体内存在不少能感受机械性刺激并引起自身功能改变的细胞。如内耳毛细胞顶部的听毛在受到切向力的作用产生弯曲时,毛细胞会出现短暂的感受器电位。这也是一种跨膜信号转换,即外来机械性信号通过某种膜结构内的过程,引起细胞的跨膜电位变化。据精细观察,从听毛受力而致听毛根部所在膜的变形到该处膜出现跨膜离子移动之间只有极短的潜伏期,因而推测可能是膜的局部变形或牵引,直接激活了附近膜中的机械门控通道。
(二)受体—G-蛋白—第二信使系统
细胞膜存在能专一性结合激素、神经递质以及其它化学物质并引起特定反应的特殊结构,称为受体(receptor) 。受体的本质是镶嵌于脂质双分子层中的大分子复合蛋白质或酶系,由结合和催化两部分组成,前者暴露在脂质双层的外表面,它的特定分子结构能够与特定的化学物质结合,好象钥匙与锁的关系。催化部分位于脂质双层的内表面,一般是一种没有活性的酶。当受体的结合部分与相应的化学物质(配体)结合时,首先引起结合部位分子构象发生变化,接着引起催化部分分子构象发生改变,使原来没有活性的酶转变成有活性酶,从而催化细胞内底物,引发一系列连锁生化反应,最后导致细胞内部功能变化。受体与配体的结合是可逆性的,并且具有特异性和饱和性的特征。实际上细胞浆和细胞核内也有受体存在,分别称为胞浆受体和核受体,但受体主要存在于膜表面。
Sutherland等人60年代在研究肾上腺素引起肝细胞中糖原分解为葡萄糖的作用机制时,发现肾上腺素只是作用于肝细胞的膜表面,通过某种发生在膜结构中的过程,先在细胞浆内生成环-磷酸腺苷(即cAMP),后者再实现肾上腺素分解糖原的作用。如果把激素这类外来化学信号看作第一信使的话,可称cAMP为第二信使(second messenger)。第二信使是由位于膜内表面的腺苷酸环化酶(AC)等膜效应器酶催化产生的。
在受体—G-蛋白—第二信使系统中,中介受体和膜效应器酶的是另一类蛋白质家族—鸟苷酸结合蛋白(guanine nucleotide-binding protein,简称G-蛋白)。已被区分出的G-蛋白有数十种,但它们的结构和功能极为相似。G-蛋白通常由α-、β-和γ-3个亚单位组成;α-亚单位通常起催化亚单位的作用,当G-蛋白与被配体激活了的受体相遇时,α-亚单位与先前结合的GDP分离而与一分子GTP结合,这时α-亚单位同其它两个亚单位分离,移动到并对邻近的膜效应器酶起作用,后者的激活(或受抑制)可以引起胞浆中第二信使物质的生成增加(或减少)(图1-16)。由于第二信使物质的生成经过多级酶催化作用,少数几个膜外化学信号分子同受体的结合,就可能在胞浆中生成数目众多的第二信使分子,这是此类跨膜信号传递的重要特点之一。
目前发现的效应器酶并不只AC一种,因而第二信使物质也不只cAMP一种,如近年来还发现,有相当数量的外界刺激信号作用于受体后,可以通过另一种G-蛋白再激活一种称为磷脂酶C的膜效应器酶,以膜结构中磷脂酰肌醇的磷脂分子为间接底物,生成两种分别称为三磷酸肌醇(IP3)和二酰甘油(DG)的第二信使,影响细胞内过程,完成跨膜信号传递。催产素、催乳素、某些下丘脑调节肽和5-羟色胺等是通过这一途径发挥作用的。可能作为第二信使的化学物质还有环-磷酸鸟苷(cGMP)、Ca2+和钙调蛋白(CaM)等。
动物体内绝大多数肽类激素、除氨基酸类以外的神经递质(约有50种)都是主要以在靶细胞中产生第二信使类物质来完成跨膜信号传递的。受体已发现100多种,一种配体往往有多种受体。受体的存在说明,引起靶细胞的生物效应不但与外来的化学物质有关,而且与细胞膜上存在的受体类型有很大关系。
上述两种主要的跨膜信号传递方式之间并不是绝对分离的,两者之间可以互相影响或在作用上有交叉。以ACh为例,当它们作用于终板膜时通过调控通道蛋白质起作用;但当ACh作用于心肌或内脏平滑肌时,遇到的却是受体—G-蛋白—第二信使系统(受体称为M-型或毒蕈碱型受体)。近年还发现,即便是M-型ACh受体,也可再区分出许多种亚型,有的亚型以cAMP为第二信使,有的以IP3和DG为第二信使。不同细胞甚或同一细胞的膜上具有对应于同一化学信号的不同受体型或其亚型,在跨膜信号传递中并不少见。
(三)酪氨酸激酶受体
近年来发现胰岛素等一些肽类激素和其他与机体发育、生长、修复、增生有关的因子,如神经生长因子、表皮生长因子、血小板源生长因子、纤维母细胞生长由子、以及与血细胞生成有关的集落刺激因子等,都是通过靶细胞表面一类称为酪氨酸激酶受体(tyrosine kinase receptor)的蛋白质起作用,这类受体结构简单,只有一个跨膜α-螺旋。当位于膜外侧的较长的肽链部分(α链)同特定的化学信号结合后,可以直接引起受体肽链的膜内段(β链)激活,使之具有磷酸激酶活性,通过使自身肽链和膜内蛋白质底物中的酪氨酸残基发生磷酸化,因而产生细胞内效应,如胰岛素作用于肌肉细胞使葡萄糖易于渗入。值得注意的是这种受体因结合配体而活化,通过自动自身磷酸化作用增大了对受体所在细胞中靶蛋白上的酪氨酸进行磷酸化的能力,并且这时即使配体从受体上解离下来,激酶的活性仍不下降(图1-17)。但是,当cAMP、IP3和DG等第二信使物质产生并发挥作用时,胰岛素受体的酪氨酸激酶的活性就大大降低。如果某个酪氨酸激酶受体只具有跨膜部分和细胞质侧部分,而缺少膜外侧与配体结合部分,就会具有永久性的酪氨酸激酶活性。失去调控序列,使它成为致癌性分子(oncogenesis)。如能引起鸟类有核红细胞增多症病毒的v-erb-B基因的表达蛋白即是。
第二章 血液
第一节血液的组成与特性
一、体液
动物体内所含的水分及其中的溶质,总称为体液(body fluid)。
细胞内液intracellular fluid):存在于细胞内的体液
体液
(60%~70%)细胞外液(extracellular fluid):存在于细胞外的体液
细胞外液可分为:分布于心血管系统内的血液(blood)中的血浆(plasma);分布于组织细胞间隙的细胞间液(interstitial fluid)或称组织液(tissue fluid)。淋巴管内的淋巴液(lymph fluid)。蛛网膜下腔、脑室、脊髓中央管内的脑脊液(cerebrospinal fluid)。也都属于组织液。细胞内液约为细胞外液的2倍。
组织液源于血液,并与细胞内液发生交换,终又回归血液。因而,血液在沟通各部位的体液,完成体内外物质交换等活动中起着尤为重要的作用。
二、血液的组成
血液由液体成分的血浆和悬浮其中的有形成分—血细胞所组成。血液的组成及主要成分所占百分比如下:
水(90%~92%)
浆(50%~60%)晶体物质(2%~3%)
白蛋白
血浆蛋白(5%~8%)球蛋白
血液(全血)纤维蛋白
红细胞
血细胞(40%~50%)白细胞
血小板
将经过抗凝处理的血液置入分血计中,经3000rpm的转速离心30分钟,血细胞则因比重较大而下沉并被压紧,可见血液分为3层:上层为微黄色或无色的液体部分即为血浆;底层为暗红色的红细胞(erythrocyte);在红细胞层表面上有一薄层灰白色物质,就是白细胞(leucocyte)和血小板(thrombocyte)。
压紧的血细胞在全血中所占的容积百分比,称为血细胞比容(hematocrit)。
在血细胞中,由于白细胞和血小板所占容积微小,可略而不计,通常就把血细胞比容称为红细胞比容,或称红细胞压积(PCV)。
血液流出血管不经抗凝处理,就会很快凝成血块,随血块逐渐紧缩所析出的淡黄色清亮液体,称为血清(serum)。
血清与血浆的主要区别在于血清中没有纤维蛋白原,因为纤维蛋白原已转变成纤维蛋白而留在了血块中。
三、血量
动物体内的血液总量,简称血量(blood volume)。
在安静状态下,大部分血液在心血管内循环流动,这部分血液量叫循环血量。另有一部分血液则滞留在肝、肺、脾、皮下静脉丛和皮肤等处,这些流动缓慢,红细胞比容较高的血液量叫储备血量或贮血量。
一次失血不超过血量的10%,一般不会影响健康,若是一次急性失血达血量的20%,生命活动将受到明显影响。倘若一次急性失血超过血量的30%,则会危及生命。
四、血液的理化特性
(一)血色、血臭、血味
血液为不透明的红色液体。动脉血中,血红蛋白氧结合量高,呈鲜红色;静脉血中,血红蛋白氧结合量低,呈暗红色。血液中由于存在挥发性脂肪酸,故带有特殊的血臭,即血腥气。又由于血液中含有氯化钠而稍带咸味。
(二)比重与红细胞的悬浮稳定性
1.比重动物全血的比重约在1.050~1.060之间,其中红细胞比重最大,白细胞和血小板次之,血浆的比重最小,约在1.025~1.030。全血比重的大小主要决定于红细胞比容的高低以及所含血红蛋白的浓度,也与血浆中血浆蛋白质的浓度有关。
2.悬浮稳定性与血沉在循环血液中,红细胞在血浆中保持悬浮状态而不易下沉的特性,称为悬浮稳定性(suspension stability)。从血管中抽取一定量的血液,经抗凝处理后放入有刻度的血沉管中,直立静置一段时间,可见红细胞因比重较大而逐渐下沉。在单位时间内红细胞下沉的距离,称为红细胞沉降率(erythrocyte sedimentation rate,ESR),简称血沉。
血沉快慢的关键,在于红细胞是否容易发生叠连。所谓叠连,系指红细胞彼此重叠在一起成串钱状。叠连起来的红细胞与血浆接触的总表面积减小,而单位面积的重量增大,于是血沉加快。凡是影响红细胞叠连的因素,就是影响血沉快慢的因素。
影响红细胞叠连的主要因素在于血浆中。已知血浆中的球蛋白,特别是纤维蛋白原与胆固醇增多时,红细胞叠连增快,血沉加速。血浆中白蛋白与卵磷脂增多时,则起延缓叠连的作用。
(三)粘滞性
液体流动时,由于内部分子间摩擦而产生阻力,以致流动缓慢并表现出粘着的特性,称为粘滞性(viscosity)。
血液粘滞性的大小,主要决定于红细胞数目的多少和血浆蛋白质的浓度。红细胞数目越多,血浆蛋白质浓度越高,血液粘滞性就越大。
粘滞性增高,血管内血流阻力增大,血流速度减慢,血压升高。
粘滞性降低,血流阻力减小,流速增快,血压降低。可见血液的粘滞性在一定程度上也是影响血压的因素之一。
(四)渗透压与红细胞的脆性
1.血浆渗透压促使纯水或低浓度溶液中的水分子通过半透膜向高浓度溶液中渗透的力量,称为渗透压(osmotic pressure)。它是晶体渗透压和胶体渗透压两部分的总和。
晶体渗透压:由血浆中的无机盐,主要是氯化钠,还有非电解质的小分子有机物,如葡萄糖和尿素等晶体物质颗粒所形成的渗透压。
胶体渗透压:由血浆蛋白质,主要是白蛋白等胶体物质颗粒所形成的渗透压。
血浆渗透压与血细胞的渗透压相等,也与机体各部位体液的渗透压基本相等。154mmol/L(0.9%〕的氯化钠溶液和278mmol/L(5%)的葡萄糖溶液的渗透压与血浆渗透压大致相等,故把154mmol/L(0.9%)的氯化钠溶液称为等渗溶液,或称为生理盐水。把那些高于或低于血浆渗透压的溶液分别称为高渗溶液或低渗溶液。
2.脆性
红细胞内血红蛋白逸出并进入血浆中的现象,称为红细胞溶解,简称溶血(hemolysis)。
红细胞在渗透压有所降低的溶液中并不一定发生溶血,说明红细胞对低渗溶液的溶解作用有一定的抵抗力。红细胞对低渗溶液的这种抵抗力,称为红细胞渗透脆性(erythrocyte osmotic fragility)或简称脆性。
(五)酸碱性
血液呈弱碱性,pH值为7.35~7.45,
血浆中的缓冲对有NaHCO3/H2CO3,蛋白质钠盐/蛋白质,Na2HPO2/NaH2PO4等;红细胞中的缓冲对有KHb/HHb,KHbO2/HHbO2等,此外,
有KHCO3/H2CO3 ,K2HPO4/KH2PO4等。
血浆中最主要的缓冲对为NaHCO3/H2CO3,当组织酸性代谢产物(如乳酸)大量进入血液时,NaHCO3即与之起作用,生成酸性较弱的碳酸和中性乳酸盐,于是酸度降低。如下列化学式所示。
HL+NaHCO3→NaL+H2CO3→H2O+CO2↑
常把血浆中NaHCO3的含量称为血液的碱储(alkali reserve)。
第二节血浆
一、血浆化学成分及其作用
(一)血浆蛋白质
1.血浆白蛋白主要由肝脏合成。其主要生理作用一是组织修补和组织生长的材料。二是形成血浆胶体渗透压。三是能与游离脂肪酸这样的脂类、类固醇激素结合,有利于这些物质的有效运输。
2.血浆球蛋白可分为α、β、γ三类,γ球蛋白几乎全都是免疫球蛋白,即抗体。除灵长类和啮齿类外。大多数新生动物只能依靠吮吸初乳来获得被动免疫。
3.纤维蛋白原由肝脏产生。是血液凝固的重要物质,在凝血酶的催化下转变为纤维蛋白,形成血凝块,起止血和凝血作用。
(二)脂类
血浆中所含的脂类包括脂肪酸、甘油三酯、磷脂、胆固醇及其它酯等。
血浆中脂类代谢的中间产物主要是酮体,血中酮体增多,大量酮体随尿排出,谓之酮尿症。
血浆中的葡萄糖称为血糖,直接氧化供给机体代谢活动所需能量,是非反刍动物体内最主要的供能物质。
(四)无机盐
重要的阳离子有Na+、K+、Ca2+等,重要的阴离子有Cl-、HCO3-、HPO42-等。这些无机离子保持一定含量,对于维持神经和肌肉组织的兴奋性、心肌细胞的活动、体液的酸碱平衡和渗透压等起着重要作用。
二、血浆的主要功能
血浆主要功能有:
1.营养功能血浆中含有相当数量蛋白质,它们起着营养贮备的功能。
2.运输功能蛋白质巨大的表面上分布有众多的亲脂性结合位点,它们可以与脂溶性物质结合,使之成为水溶性,便于运输
3.缓冲功能血浆白蛋白和它的钠盐组成缓冲对,和其它无机盐缓冲对(主要是碳酸和碳酸氢钠)一起,缓冲血浆中可能发生的酸碱变比,保持血液pH的稳定,
4.形成胶体渗透压血浆胶体渗透压的存在是保证血浆中的水分不会大量向血管外转移的重要条件,从而维持血量的相对恒定。
5.参与机体的免疫功能在实现免疫功能中有重要作用的免疫抗体、补体系统等,都是由血浆球蛋白构成的。
6.参与凝血和抗凝血功能绝大多数的血浆凝血因子、生理性抗凝物质以及促进血纤维溶解的物质都是血浆蛋白。
7.组织生长与损伤组织修复方面的功能这是由白蛋白转变为组织蛋白而实现的。
第三节红细胞
一、红细胞的形态和数目
(一)形态
哺乳动物的红细胞是无核、双凹圆盘形细胞,驼骆和鹿的红细胞呈椭圆形。双凹圆盘形细胞容易变形。
(二)数目
常以1012个/l血液表示。
二、红细胞的功能与血红蛋白
红细胞的主要功能是运输O2和CO2,这项功能是由红细胞所含的血红蛋白(haemoglobin,Hb)来完成的。
HB是由珠蛋白与亚铁血红素组成的结合蛋白质。血红蛋白容易与氧疏松结合成氧合血红蛋白,在氧分压低时,氧又容易解离而释放出来,血红蛋白也能与二氧化碳结合成氨基甲酸血红蛋白,或称碳酸血红蛋白,在CO2分压低的环境中,CO2又解离释放出来,此外,血红蛋白及其钾盐构成的缓冲对是调节血液酸碱平衡的重要缓冲物质。
在正常情况下,单位容积内红细胞数目与血红蛋白含量的高低是基本一致的。如果红细胞数目和血红蛋白含量都减少,或其中之一明显减少,都可视为贫血。
血红蛋白与CO的亲合力比对O2的亲合力大200余倍,并结合成稳定的一氧化碳血红蛋白。
三、红细胞的生成与破坏
(一)红细胞的生成
1、生成动物出生后,红细胞由红骨髓生成。
2.生成的条件
造血原料以蛋白质、二价铁离子最为重要, 此外,还需要维生素B6 、B2、C、E,微量元素铜、锰、钴和锌等。
3.红细胞生成的调节
主要受促红细胞生成素(erythropoietin, EPO)的调节,雄激素等也参与其调节过程。
红细胞在体内破坏增多时,从细胞内释放出来的产物也能刺激红细胞生成加速。
(二)红细胞的破坏
1、红细胞主要是因衰老而遭破坏。衰老红细胞的变形能力减退,脆性增大,因而易受血液的机械冲击而破损。2、容易被吞噬细胞所吞噬。
第四节 白细胞
- 白细胞的形态与数目
(一)形态
白细胞是比红细胞体积大、数目少、比重小的有核血细胞。
分类:中性粒细胞
粒性细胞嗜酸性粒细胞
嗜碱性粒细胞
白细胞淋巴细胞
无颗粒细胞
单核细胞
白细胞与红细胞和血小板一样都起源于骨髓中的造血干细胞。
(二)数目
常以109个/升血液计数。
二.白细胞的免疫功能
依据免疫功能的特点,可将白细胞分为吞噬细胞和免疫细胞两大类。吞噬细胞(phagocyte)主要指中性粒细胞和单核细胞;免疫细胞主要指淋巴细胞。吞噬细胞主要通过吞噬作用来消灭异物,不具有针对某一异物的特异性,故称为非特异性免疫。免疫细胞消灭异物的主要方式是产生抗体和局部细胞反应,都具有针对性很强的特异性,所以称为特异性免疫。
(一)中性粒细胞
中性粒细胞吞噬和作变形运动的能力都很强,又具有复杂的杀菌系统。
(二)嗜碱性粒细胞
嗜碱性粒细胞主要在组织中发挥作用,但缺乏吞噬能力,其结构与功能都与结缔组织中的肥大细胞相似。能释放组胺、过敏性慢作用物质、嗜酸性粒细胞趋化因子A、肝素等活性物质。
(三)嗜酸性粒细胞
具有变形运动能力,它虽含有溶酶体,但缺乏溶菌酶,基本上不具杀菌能力,溶酶体中含有一种能沉淀DNA、中和肝素的碱性蛋白质。
(四)单核—巨噬细胞
单核细胞能分裂增殖,能作变形运动,但吞噬能力很弱。单核细胞在血流中停留2~4d后,随即进入肝、脾、肺、淋巴结和浆膜腔等部位,转变成巨噬细胞。巨噬细胞的特点是体积增大,溶酶体和溶菌酶增多,吞噬能力大为增强,惟增殖能力丧失。因此将二者合称为单核—巨噬系统,是机体内一个庞大的防御系统。
(五)淋巴细胞
淋巴细胞是具有特异性免疫功能的免疫细胞,它们在免疫应答过程中起着核心作用。
1.T细胞
主要功能是实施细胞免疫。
2.B细胞
主要功能是实现体液免疫。
三、白细胞的生成和破坏
(一)白细胞的生成
一般在骨髓颗粒细胞,但淋巴细胞除来自骨髓外,还在淋巴组织包括胸腺、淋巴结、扁桃体等产生;单核细胞部分在骨髓,部分在网状内皮系统产生。
(二)白细胞的破坏可因衰老死亡和执行防御功能被消耗而致。有的被网状内皮系统吞噬,有的则通过消化、呼吸、泌尿道排出体外。
第五节生理止血机制
一、血小板
血小板(platelets thrombocyte)是从骨髓成熟的巨核细胞胞浆裂解脱落下来的小块胞质。
(一)血小板的形态、数目
循环血液中的血小板是无色透明、无细胞核、圆盘形或杆形小体。
(二)血小板的生理特性
1.粘附与聚集粘附系指血小板易于附着在异物表面。当血管内皮损伤,暴露出内皮下的胶原纤维时,使血小板激活并粘附其上。
所谓聚集,是指血小板与血小板相互粘着在一起,聚集成团的现象。
2.吸附与释放血小板能吸附血浆中的凝血因子(blood coagulation factor),使血小板聚集的局部凝血因子的浓度增高,促进凝血反应,血小板还能从血浆中主动吸收5-羟色胺(5-HT)、儿茶酚胺等,贮存于致密颗粒中。
血小板激活后、可将贮存颗粒中的ADP、5-HT和儿茶酚胺等活性物质释放出来。内源性的ADP又进一步使血小板聚集,5-HT和儿茶酚胺使小动脉收缩,均有利于止血。
3.收缩血小板内含有血小板收缩蛋白,使血小板具有收缩性,可促使凝血块紧缩、止血栓硬化,加强止血效果。
(三)血小板的功能
血小板的主要功能是参与止血(hemostasis)和加速血液凝固(blood coaguation)。
1.止血功能
2.凝血功能
3.对纤维蛋白溶解的作用
4.营养与支持作用
(四)血小板的生成与破坏
1.血小板的生成 生成血小板的巨核细胞也是从骨髓中的造血干细胞分化发展来的。
2.血小板的破坏血小板生存时间很短,在循环血液中平均寿命只7~14天,并且只有在进入血液后的最初2天具有生理功能。血小板可因衰老而遭破坏或在发挥生理功能中被消耗,衰老的血小板绝大部分是在脾、肝、肺和骨髓内被网状内皮细胞所吞噬。
二、血液凝固
血液离开血管后,很快就由流动的溶胶状态变成不流动的胶冻状凝块,这一过程称为血液凝固(blood coagulation),简称血凝。
(一)凝血因子
血浆与组织直接参与凝血的物质,统称为凝血因子(blood clotting factors)
(二)凝血过程
凝血过程基本上是一系列蛋白质有限水解的过程,凝血过程一旦开始,各个凝血因子便一个激活另一个,形成一个“瀑布”样的反应链直至血液凝固。凝血过程大致可分为三个阶段(图2-3)。
第一阶段:凝血因子X激活成Xa(活化型),并形成凝血酶原酶复合物;
第二阶段:凝血酶原(Prothrombin)(因子Ⅱ)激活成凝血酶(thrombin)(Ⅱa);
第三阶段:纤维蛋白原(因子Ⅰ)转变成纤维蛋白(fibrin)(Ⅰa)。
三、抗凝血系统与纤维蛋白溶解
(一)血液中的抗凝系统
血液在心血管系统内循环,之所以不发生凝固一方面是因为血管内膜光滑,不出现异物面,凝血因子Ⅻ不可能发生表面激活,血小板的粘附、聚集也不能发生。再则,即使血浆中有少量凝血因子成了活化型,也将被稀释,不足以引起凝血反应,并由肝脏清除或被吞噬细胞吞噬。更重要的一个方面是血浆中存在着一些抗凝物质。通常把这些抗凝物质统称为抗凝系统。
现已查明,血浆中最重要的抗凝物质是抗凝血酶Ⅲ(antithrombin Ⅲ)和肝素,它们的作用约占血浆全部抗凝血酶活性的75%。抗凝血酶Ⅲ是血浆中一种丝氨酸蛋白酶抑制物(serine protease inhibitor)。因子Ⅱa、Ⅶ、Ⅸa、Ⅹa、Ⅻa 的活性中心均含有丝氨酸残基,都属于丝氨酸蛋白酶(serine protease)。抗凝血酶 Ⅲ分子上的精氨酸残基,可以与这些酶活性中心的丝氨酸残基结合,这样就“封闭”了这些酶的活性中心而使之失活。
肝素是一种酸性粘多糖,主要由肥大细胞和嗜碱性粒细胞产生,存在于大多数组织中,在肝、肺、心和肌组织中更为丰富。
肝素在体内和体外都具有抗凝作用倍。
(二)纤维蛋白溶解
凝血过程中形成的纤维蛋白,在一定条件下又可以发生溶解的过程,称为纤维蛋白溶解(fibrinolysis),简称纤溶。
1.纤溶酶原的激活 纤溶酶原很可能是在肝、骨髓、嗜酸性粒细胞和肾组织中合成,在激活物的作用下成为纤溶酶。
2.纤维蛋白与纤维蛋白原的降解纤溶酶是血浆中特异性很小而活性甚强的蛋白酶。它的主要作用是水解纤维蛋白原和纤维蛋白。除此之外还能水解其它一些蛋白质,如凝血酶、凝血因子V和Ⅷ;激活因子Ⅻa;也能促进血小板的聚集和释放5-羟色胺,ADP等;还能激活血浆中的补体系统。纤溶酶对纤维蛋白的作用是水解纤维蛋白与纤维蛋白原分子中肽链上各部位的赖氨酸—精氨酸键,从而将纤维蛋白与纤维蛋白原分子分割成很多可溶性的小肽。这些可溶性的小肽,总称为纤维蛋白降解产物。纤维蛋白降解产物一般不能再出现凝固,其中一部分有抗血凝作用。
3.抑制物及其作用血浆中存在的纤溶抑制物主要是抗纤溶酶(antiplasmin),其特异性也不大。
四、促进和延缓血液凝固
(一)促凝
1.机体受创伤出血时,使血液与粗糙面接触,这样既可促进凝血因子Ⅻ的激活,又可促进血小板聚集、解体并释放凝血因子,加速凝血反应的进程。提高创口部位的温度可以加快酶促反应的速度,也能加速血凝。所以,临床上用棉花球、明胶海绵、温热生理盐水浸渍的纱布按压创口,可收到良好的止血效果。
2.维生素K在肝脏内参与凝血酶原和凝血因子Ⅶ、Ⅸ、Ⅹ等的合成过程,有加速凝血和止血的间接作用。
(二)抗凝
1.移除血浆中的钙离子
2.去除纤维蛋白
3.低温延缓血凝较低的温度可使凝血过程中酶促反应减慢,使血凝延缓。
4.血液与光滑面接触如容器内预先涂一层石蜡,可因凝血因子Ⅻ的活化延迟等原因而使血凝延缓。
5.肝素在有抗凝血酶Ⅲ存在时,肝素对凝血过程各阶段都有抑制作用。无论在体内或体外它都是很强的抗凝剂(anticoagulation),并具有用量少,对血液影响小,保存性好等优点。
6.双香豆素
第六节血型
一、有关血型的几个概念
(一)血型
血型(blood group)通常指红细胞膜上特异性抗原的类型。广义的血型定义是指由遗传决定的血液、脏器以及分泌物的各种特性。主要有两大类:一类是以细胞膜抗原结构的差异为特征的血细胞抗原型,即能用抗体进行分类的血细胞抗原型;另一类是以蛋白质化学结构微小差异为特征的蛋白质多态性和同工酶,即通过一定的方法能区分出的各种蛋白质和酶的遗传变异型,也即所谓的蛋白质型和酶型。
(二)血型系统与血型因子
由同一遗传位点等位基因控制的血型系列,称为血型系统。血型抗原是指由遗传决定的、具有抗原特性的特殊结构。一种血型抗原可能有一种以上的抗原特异性,引起几种不同的抗体产生。在血型上,引起不同抗体产生并能与之发生反应的抗原成分称为血型因子,又称为抗原因子。血型因子通常用正体的大写英文字母,或在大写英文字母后标一小写字母来表示。
(三)基因型与表现型
基因型即基因的组合,它是决定个体特性的基因型式。表现型是与基因型相对应的实际表现出来的性状。血型即是与基因型相对应的实际表现出来的性状,由基因型决定。
二、红细胞血型
通常所说的血型是指红细胞的血型,但是存在于红细胞上的血型抗原也存在于白细胞、血小板和一般组织细胞上。此外,在白细胞和血小板上还存在它们本身特有的抗原。
(一)人类的ABO血型系统
人类红细胞膜上含A凝集原(A抗原)和B凝集原(B抗原)两种抗原物质,血清中含α凝集素(抗A)和β凝集素(抗B)两种抗体。凡红细胞膜上只含A抗原者为A型,红细胞膜上只含B抗原的,称为B型;若A与B两种抗原都含有的称为AB型;这两种抗原都没有的,则称为O型。不同血的人的血清中含不同的凝集素,即不含有对抗他自身红细胞凝集原的凝集素。在A型人的血清中,只含有抗B凝集素;B型人的血清中,只含有抗A凝集素;AB型人的血清中没有抗A和抗B的凝集素;而O型血人的血清中则含有抗A和抗B两种凝集素(表2-9)
(二)Rh血型系统
人类在寻找新血型物质的探索中,把恒河猴的红细胞重复注射入家兔体内,引起家兔产生免疫反应,使家兔的血清中产生抗恒河猴红细胞的抗体(凝集素)。再用含这种抗体的血清与人的红细胞混合,发生血清凝集反应者称为Rh阳性血型,表明其红细胞上具有与恒河猴同样的抗原;若人的红细胞不被这种血清凝集,称为Rh阴性血型。
(三)家畜的血型
家畜血清中天然存在的红细胞血型抗体已知的还不太多,而且免疫效价也很低,所以同种个体间首次输血,一般不会引起严重后果。但再次输血时就应该先作交叉配血试验,即把供血者与受血者的红细胞分别用对方的血清来作凝集反应,并判断是否可以输血。
蛋白质型指同种不同个体中,具有相同功能的蛋白质所存在的多态性。酶蛋白的多态性称为同功酶。这样的多态性不仅血液蛋白中存在,乳汁、精液等分泌物以及脏器中也存在,是广义的血型概念。蛋白质分子中分子结构与分子大小不同,可用电泳法将它们分离出来,显现出受遗传控制的一些区带,以此划分蛋白质类型。
蛋白质型与红细胞血型的记载方法不尽相同。如运铁蛋白型,记为Tf型,电泳分离出来的A带、B带等,记为TfA、TfB,其等位基因记为TfA、TfB。
常有报道的家畜蛋白质型和酶型有:白蛋白型(Alb型)、前白蛋白型(Pr型)、后白蛋白型(Pa型)、运铁蛋白型(Tf型)、血浆铜蓝蛋白型(Cp型)、血液结合素型(HP型)、血浆脂蛋白型(Lpp型)、血红蛋白型(Hb型)、碳酸酐酶型(AC型)、淀粉酶型(Am型)、碱性磷酸酶型(AKP型)、脂酶型(ES型)、6-磷酸葡萄糖脱氢酶型(6-PGD型)、乳酸脱氢酶型(LDH型)等等。
三、血型的应用
血型除在输血时有重要意义外,在畜牧和兽医实践中均有广泛应用。
(一)血统登记和亲子鉴定
血型是完全遗传而又终生不变的,通过血型登记,记载能稳定遗传给后代的血型,从而把祖先和后代的登记联系起来,以建立准确的系谱资料,防止血统混乱,保证育种工作的可靠性。
(二)诊断异性孪生不育
牛怀孕异性双胎时,两胎儿间血管吻合发生率约有12%。在发生血管吻合的情况下,一方面雄性胎儿性腺产生的雄激素可作用于尚未分化的雌性胎儿性腺,影响雌性胎儿性腺的分化,使产出的母犊日后缺乏生殖能力;另一方面,胎儿血管吻合后,造血器官中
的原红细胞能进入对方体内。这样使胎儿具有两种红细胞,这一现象叫红细胞嵌合。
(三)血型与新生仔畜溶血病
母子间血型不合,胎儿的血型抗原物质进入母体后,引起母体产生血型抗体,这种抗体不能通过胎盘,只能在分娩之后经初乳转移给仔畜,造成仔畜的红细胞迅速破坏而发生溶血,呈急性溶血和黄疸症状,且能致死。
(四)组织相容性
异体器官或组织能相处并发挥正常功能的能力,称为相容性。由于存在免疫反应,机体往往对异体器官表现排斥反应。白细胞,特别是淋巴细胞血型所表现的相容性,能在一定程度上反映组织移植的相容性。因此,常将受体与供体的淋巴细胞混合作组织培养,根据细胞分裂状态来判别两者不相容的程度。随着组织细胞相容性抗原的研究不断深入,将有力推动器官移植工作的发展。
(五)利用血型研究物种起源与亲缘关系
用一种动物的免疫血清检测不同动物的红细胞凝集反应,反应越强的动物,表明与该种动物的亲缘关系越亲近,反应越弱的则与该种动物的亲缘关系越疏远。
(六)血型与生产性能的关系
血型是动物的一种遗传性状,而动物的各种经济性状在某种程度上是受遗传控制所表现的遗传性状,控制动物这两类遗传性状的基因可能存在某种直接或间接的联系。这样就激起了许多学者探索血型与生产力、抗病力等相关性研究的极大兴趣,力图查明这种关系,以作为优良个体选育和品种改良的可靠依据。已有红细胞型与奶牛产奶量性能的关系之研究,运铁蛋白型与乳脂率、繁殖率的关系之研究等诸多有价值的研究报导。迄今,以上方面的研究仍十分活跃。
第三章 神经系统
第一节神经元活动的一般规律
神经元是神经系统的结构和功能单位,
神经元
1、细胞体
2、突起
树突:短而粗,分支多
轴突:细而长,即通常所说的神经纤维
一、神 经 纤 维
(一)神经纤维传导的特征
1.生理完整性神经纤维被切断后,破坏了结构上的完整性,冲动就不能传导。如果结扎或在麻醉药、低温等作用下,使神经纤维机能发生改变,破坏了生理功能的完整性,冲动传导也将发生阻滞。
2.绝缘性一条神经干内有许多神经纤维,其中包含有传入和传出纤维,各条纤维上传导的兴奋基本上互不干扰,准确地实现各自的功能,这种特点叫做绝缘性传导。
3.双向性刺激神经纤维上的任何一点,兴奋就从刺激的部位开始沿着纤维向两端传导,叫做传导的双向性。
4.不衰减性神经纤维在传导冲动时,不论传导距离多长,其冲动的大小,频率和速度始终不变,这一特点称为传导的不衰减性。这对于保证及时、迅速和准确地完成正常的神经调节功能十分重要。
5.相对不疲劳性在实验条件下,用每秒50~100次的电刺激连续刺激蛙的神经9~12h冲动仍能传导,这说明神经纤维是不容易发生疲劳的。
(二)神经纤维的分类
1.根据电生理学的特性分类
A类:包括有髓的躯体传入和传出纤维,依据其平均传导速度,又进一步分为α、β、γ、δ四类。
B类:有髓的植物性神经的节前纤维。
C类:包括无髓的躯体传入纤维(dγC)和植物性神经节后纤维(SC)。
2.根据纤维直径的大小和来源分类将传入纤维分为Ⅰ、Ⅱ、Ⅲ、Ⅳ四类
(三)神经纤维的传导速度
1、与纤维直径有关:神经纤维越粗,传导速度越快。有髓纤维的传导速度和直径成正比,传导速度(m/s)= 6×直径(μm)。
2、与温度有关:温度降低则传导速度减慢。
二、突触
两个神经元相接触的部位就称之为突触(synapse)。在突触前面的神经元叫突触前神经元,在突触后面的神经元叫突触后神经元。
(一)突触的分类
1.根据突触接触部位分类
(1)轴—树突触(axo-dendritic synapse)指神经元的轴突末梢与下一个神经元的树突发生接触。
(2)轴—体突触(axo-somatic synapse)指一个神经元的轴突末梢与下一个神经元的胞体发生接触。
(3)轴—轴突触(axo-axonic synapse)指一个神经元的轴突末梢与下一个神经元的轴丘(轴突始段)或轴突末梢发生接触。
此外,还存在树—树、体—体、体—树及树—体等多种形式的突触联系。近年来还发现,同一个神经元的突起之间还能形成自身突触(autoapse)。
2.按突触性质分类
a化学性突触
(1)兴奋性突触(excitatory synapse)
(2)抑制性突触 (inhibitory synapse)
b电突触
(二)突触的基本结构
1.化学性突触(chemical synapse)
概念:
(1)突触小体:一个神经元的轴突末梢首先分成许多小支,每个小支的末端膨大呈球状,称突触小体。
(2)突触小泡:在突触小体内含有较多的线粒体和大量的小泡,此小泡称为突触小泡。内含有兴奋性递质或抑制性递质。突触小泡线粒体内含有合成递质的酶。
(3)突触间隙:小体与另一神经的胞体或树突形成突触联系。称为突触间隙。
(4)突触前膜:由突触小体构成突触间隙的膜称突触前膜。
(5)突触后膜:构成突触间隙的另一侧膜称突触后膜。上有特殊的受体,能与专一的递质发生特异性结合。
一个突触即由突触前膜、突触间隙和突触后膜3部分构成。
2.电突触(electrical synapse)
电突触的结构基础是缝隙连接,其突触前神经元的轴突末梢内无突触小泡,也无神经递质。连接部位存在沟通两细胞胞浆的通道,带电离子可通过这些通道而传递电信号,因此这种连接部位的信息传递是一种电传递。
(三)突触传递
冲动从一个神经元通过突触传递到另一个神经元的过程,叫做突触传递(synaptic transmission)。
- 化学性突触的传递
(1)神经冲动传至轴突末梢时,突触前膜兴奋,爆发动作电位和离子转移。此时突触前膜对Ca2+的通透性加大,Ca2+由突触间隙顺浓度梯度流入突触小体,然后小泡内所含的化学递质以量子式释放的形式释放出来,到达突触间隙。
(2)递质释放出来后,通过突触间隙,扩散到突触后膜,与后膜上的特殊受体结合,改变后膜对离子的通透性,使后膜电位发生变化。这种后膜的电位变化,称为突触后电位(postsynaptic potential)。突触后电位有两种类型,即兴奋性突触后电位和抑制性突触后电位。
(1)兴奋性突触后电位当动作电位传至轴突末梢时,使突触前膜兴奋,并释放兴奋性化学递质(excitatory transmitter),递质经突触间隙扩散到突触后膜,与后膜的受体结合,使后膜对Na+、K+、Cl-,尤其是对Na+的通透性升高,Na+内流,使后膜出现局部去极化,这种局部电位变化,叫做兴奋性突触后电位(excitatory postsynaptic potential, EPSP)。此过程称兴奋性突触传递。
(2)抑制性突触后电位当抑制性中间神经元兴奋时,其末梢释放抑制性化学递质(inhibitory transmitter)。递质扩散到后膜与后膜上的受体结合,使后膜对K+、Cl-,尤其是对Cl-的通透性升高,K+外流和Cl-内流,使后膜两侧的极化加深,即呈现超极化,此超极化电位叫做抑制性突触后电位(inhibitory postsynaptic potential, IPSP),此过程称抑制性突触传递。
化学性突触的传递,可概括如下:
突触前神经元末梢兴奋→释放兴奋性递质→兴奋性突触后电位(突触后膜去极化)→突触后神经元兴奋。突触前神经元末梢兴奋→释放抑制性递质→抑制性突触后电位(突触后膜超极化)→突触后神经元抑制。
2.电突触的传递
突触前神经元的动作电位到达神经末梢时,引起突触后成分发生动作电位。在冲动未到达突触前末梢时,对突触后膜有阳极电紧张作用,使突触后膜的膜电位升高、兴奋性降低。当动作电位传到突触前末梢时,神经末梢呈负性,就好像一个阴极,起阴极电紧张作用,使突触后膜的膜电位下降,兴奋性提高。当兴奋性提高到一定程度时,就产生神经冲动,并以局部电流进行传播。
3.非突触性化学传递
肾上腺素能神经元的轴突末梢有许多分支,在分支上有大量结节状曲张体,曲张体内含有大量的小泡,是递质释放的部位。当神经冲动抵达曲张体时,递质从曲张体释放出来,通过弥散作用到达效应器细胞的受体,使效应细胞发生反应。由于这种化学传递不是通过突触进行的,故称为非突触性化学传递。
非突触性化学传递与突触性化学传递相比,有下列几个特点:
①不存在突触前膜与突触后膜的特化结构。
②不存在一对一的支配关系,即一个曲张体能支配较多的效应细胞。
③曲张体与效应细胞间的距离至少在200?以上,距离大的可达几个μm。
④递质的弥散距离大,因此传递花费的时间可大于1s。
⑤递质弥散到效应细胞时,能否发生传递效应取决于效应细胞膜上有无相应的受体存在。
三、神 经 递 质
(一)外周神经递质
1.乙酰胆碱植物性神经全部节前纤维、副交感神经节后纤维、部分交感神经节后纤维(支配汗腺和骨骼肌血管等交感神经节后纤维)和躯体运动神经纤维都是释放乙酰胆碱作为递质的。凡是释放乙酰胆碱作为递质的神经纤维,均称为胆碱能纤维(cholinergic fibres)。植物神经节前纤维和运动神经纤维所释放的乙酰胆碱的作用,与烟碱的药理作用相同,称为烟碱样作用(N样作用);而副交感神经节后纤维所释放的乙酰胆碱的作用,与毒蕈碱的药理作用相同,称为毒蕈碱样作用(M样作用)。
2.去甲肾上腺素在动物机体中绝大多数交感神经节后纤维是释放去甲肾上腺素作为递质的。凡是释放去甲肾上腺素作为递质的神经纤维,称为肾上腺素能纤维(adrenergic fibres)。在高等动物中由交感神经节后纤维释放的递质仅是去甲肾上腺素,而不含肾上腺素。
3.嘌呤类和肽类递质植物神经的节后纤维除胆碱能和肾上腺素能纤维外,还有第三类纤维。第三类纤维末梢释放的递质是嘌呤类和肽类化学物质。
(二)中枢神经递质
1.乙酰胆碱乙酰胆碱是中枢神经系统的重要递质。如脊髓腹角运动神经元、脑干网状结构的前行激动系统、纹状体(尤其是尾状核)内部具有乙酰胆碱递质。乙酰胆碱在这些地方一般起兴奋性递质的作用。
2.单胺类单胺类包括多巴胺(dopamine)、去甲肾上腺素和5-羟色胺。
多巴胺主要由黑质制造,沿黑质—纹状体系统分布,在纹状体内贮存,是锥体外系统的重要递质,它与躯体运动协调机能有关,一般起抑制性作用。
a、中脑网状结构的N元与躯体运动和内脏活动调节有关
产生部位b、脑桥的蓝斑核的N元与维持醒觉有关
c、延髓网状结构的N元与情绪反应和下丘脑内分泌调节功能有关
5-羟色胺主要由脑干背侧正中线附近的中缝核群产生。其纤维向前投射到纹状体、丘脑、下丘脑、边缘系统和大脑皮质,与睡眠、情绪反应、调节下丘脑的内分泌功能有关;后行纤维到达脊髓,与躯体运动和内脏活动的调节有关。
3.氨基酸类氨基酸类包括谷氨酸、甘氨酸和γ氨基丁酸等。谷氨酸是兴奋性递质,广泛分布于大脑皮质和脊髓内,与感觉冲动的传递及大脑皮质内的兴奋有关,甘氨酸在脊髓腹角的闰绍细胞浓度最高,引起突触后膜超极化,产生突触后抑制。γ氨基丁酸在大脑皮质的浅层和小脑的浦肯野氏细胞含量较高,引起突触后膜超极化,产生突触后抑制。γ氨基丁酸在脊髓内能引起突触前膜去极化,产生突触前抑制。
4.肽类视上核和室旁核神经元分泌升压素(9肽)和催产素(9肽);下丘脑内其他肽能神经元能分泌多种调节腺垂体活动的多肽,如促甲状腺激素释放激素(TRH,3肽)、促性腺激素释放激素(GnRH,10肽)、生长抑素(GHRIH,14肽)等。由于这些肽类物质在分泌后,要通过血液循环才能作用于效应细胞,因此称为神经激素。但现已知,这些肽类物质可能还是神经递质。
脑内具有吗啡样活性的多肽,称为阿片样肽。阿片样肽包括β-内啡肽、脑啡肽和强啡肽三类。脑啡肽是5肽化合物,有甲硫氨酸脑啡肽(M-ENK)和亮氨酸脑啡肽(L-ENK)两种。脑啡肽与阿片受体常相伴而存在,微电泳脑啡肽可使大脑皮层、纹状体和中脑导水管周围灰质神经元的放电受到抑制。脑啡肽在脊髓背角胶质区浓度很高,它可能是调节痛觉纤维传入活动的神经递质。
此外,脑内还有其他肽类物质,如P物质、神经降压素、血管紧张素Ⅱ、胆囊收缩素(CCK)、促胰液素、胃泌素、胃动素、血管活性肠肽、胰高血糖素等。
5.其他可能的递质近年来研究指出,一氧化氮具有许多神经递质的特征。某些神经元含有一氧化氮合成酶,该酶能使精氨酸生成一氧化氮。生成的一氧化氮从一个神经元弥散到另一神经元中,而后作用于鸟苷酸环化酶并提高其活力,从而发挥出生理作用。此外,组胺也可能是脑内的神经递质。
(三)递质与调质的概念
递质是指神经末梢释放的特殊化学物质,它能作用于支配的神经元或效应器细胞膜上的受体,从而完成信息传递功能。
调质是指神经元产生的另一类化学物质,它能调节信息传递的效率,增强或削弱递质的效应。
(四)递质的共存
在无脊椎动物的神经元中,观察到多巴胺和5-羟色胺递质可以共存。在高等动物的交感神经节神经元发育过程中,去甲肾上腺素和乙酰胆碱可以共存。此外,在大鼠延髓的神经元中观察到5-羟色胺和P物质共存;在上颈交感神经节神经元中观察到去甲肾上腺素和脑啡肽共存,
(五)受体学说
目前认为神经递质,必须先与突触后膜或效应器细胞上的受体相结合,才能发挥它的作用。如果受体事先被某种药物结合,则递质很难再与受体结合,于是递质就不能发挥其作用。这种与受体结合使递质不能发挥作用的药物,叫做受体阻断剂(antogonism)。递质与其相应的受体阻断剂在化学结构上往往具有一定的相似性,因此两者均能和同一受体结合并发生竞争。
1.胆碱能受体凡是能与乙酰胆碱结合的受体叫做胆碱能受体(cholinoceptor)。胆碱能受体又可分为两种:一种是毒蕈碱型受体(muscarinic receptor)或M受体,它与乙酰胆碱结合时产生与毒蕈碱相似的作用。另一种叫烟碱型受体(nicotinic receptor)或N受体,它与乙酰胆碱结合时产生与烟碱相似的作用。
M型受体存在于副交感神经节后纤维支配的效应细胞上以及交感神经支配的小汗腺、骨骼肌血管壁上。当它与乙酰胆碱结合时,则产生毒蕈碱样作用,也就是使心脏活动受抑制、支气管平滑肌收缩、胃肠运动加强、膀胱壁收缩、瞳孔括约肌收缩、消化腺及小汗腺分泌增加等。阿托品可与M受体结合,阻断乙酰胆碱的毒蕈碱样作用,故阿托品是M受体的阻断剂。
N受体又可分为神经肌肉接头和神经节两种亚型,它们分别存在于神经肌肉接头的后膜(终板膜)和交感神经、副交感神经节的突触后膜上,前者为N2,后者为N1受体类型。当它们与乙酰胆碱结合时,则产生烟碱样作用,即可引起骨骼肌和节后神经元兴奋。箭毒可与神经肌肉接头处的N2受体结合而起阻断剂的作用;六烃季胺可与交感、副交感神经节突触后膜上的N1受体结合而起阻断剂的作用。
2.肾上腺素能受体(adrenoceptor)凡是能与儿茶酚胺(catecholamine)(包括去甲肾上腺素、肾上腺素等)结合的受体称之为肾上腺素能受体。其对效应器的作用,有兴奋效应也有抑制效应。肾上腺素能受体又可分为α和β两种。α受体与儿茶酚胺结合后,主要是兴奋平滑肌,如血管收缩、子宫收缩和瞳孔开张肌收缩等;但也有抑制作用,如使小肠平滑肌舒张。β受体又可分为β1和β2两个亚型,它与儿茶酚胺结合后,抑制平滑肌的活动,如血管舒张、子宫收缩减弱、小肠及支气管平滑肌舒张等,但对心肌却产生兴奋效应。一般说来,递质与α受体结合后引起效应器细胞膜的去极化,而与β受体结合后则引起超极化,因而出现不同的效应。
3.突触前受体近年来的研究指出,受体不仅存在于突触后膜,而且也存在于突触前膜。突触前膜的受体叫做突触前受体。突触前受体的作用,在于调节神经末梢的递质释放。例如,肾上腺素能纤维末梢的突触前膜上存在α受体,当末梢释放的去甲肾上腺素在突触前膜处超过一定量时,即可与突触前膜α受体结合,从而反馈抑制末梢合成和释放去甲肾上腺素,起到调节末梢递质释放量的作用。
4.中枢内递质的受体中枢递质种类复杂,因此相应的受体也多,除胆碱能N型和M型受体、肾上腺素能α和β受体外,还有多巴胺受体、5-羟色胺受体、兴奋性氨基酸受体、γ氨基丁酸受体、甘氨酸受体、阿片受体(opiate receptor)等。
第二节 反射中枢活动的一般规律
- 中 枢 兴 奋
中枢兴奋的传布有以下特征:
1.单向传布在中枢神经系统中,冲动只能沿着特定的方向和途径传播,即感受器兴奋产生的冲动向中枢传递,中枢的冲动则传向效应器,这种现象称为单向传布。单向传布是由突触传递的特征决定的。
2.反射时和中枢延搁从刺激作用于感受器起,到效应器发生反应所经历的时间,称为反射时(reflex time),这是兴奋通过反射弧(relex arc)各个环节所需的时间。其中兴奋通过突触时,经历时间较长,即所谓突触延搁,在突触传递过程中,必须经历化学递质的释放、扩散、与后膜上的受体结合,产生兴奋性突触后电位,再通过总和作用,才使突触后神经元兴奋,故延搁时间较长。这种现象称为中枢延搁(central delay)。由于中枢延搁和突触存在有关,因此在中枢内的突触联系越多,反射时就越长。
3.总和(summation)在突触传递中,如果同一突触前末梢连续传来多个冲动,或多个突触前末梢同时传来一排冲动,则突触后神经元可将所产生的突触后电位总和起来,待达到阈电位水平时,就使突触后神经元兴奋,前者称为时间总和,后者称为空间总和。在中枢内兴奋的总和实际上就是突触总和。
4.扩散与集中集中:如,饲喂时,由嗅觉、视觉和听觉器官传入中枢的冲动,可共同引起唾液分泌中枢的兴奋,从而导致唾液分泌。兴奋集中的结构基础是由于中枢内的神经元存在着聚合式突触联系的结果。上述兴奋的空间总和即兴奋集中的表现。
扩散(diffusion)。如,当皮肤受到强烈的伤害性刺激时,所产生的兴奋传到中枢后,在引起机体的许多骨骼肌发生防御性收缩反应的同时,还出现心血管、呼吸、消化和排泄系统等活动的改变,这就是中枢兴奋扩散的结果。
5.兴奋节律的改变传出神经的兴奋除取决于传入冲动的节律外,还取决于传出神经元本身的功能状态。在多突触反射中则情况更复杂,冲动由传入神经进入中枢后,要经过中间神经元的传递,因此传出神经元发放的频率还取决于中间神经元的功能状态和联系方式。
6.后放在一个反射活动中,常可看到,当刺激停止后,传出神经仍可在一定时间内连续发放冲动,使反射能延续一段时间,这种现象称为后放(after discharge)。
7.易化作用和抑制作用
易化作用:中枢内每一神经元兴奋性可受到其它神经元的影响而发生变化。当其兴奋性受到影响而升高时,其兴奋阈值降低,则兴奋的传递易于进行,反射易于发生,这一现象称为中枢兴奋的易化作用。
抑制作用:当某一神经元的兴奋性因受到其它神经元的影响而降低时,则兴奋阈值就升高,使中枢兴奋的传递难以进行,反射也较难发生,这一现象称为中枢兴奋的抑制作用。
8.对内环境变化的敏感性和易疲劳性突触是反射弧中最易发生疲劳的部位。突触也最易受内环境变化的影响。突触对某些药物亦很敏感。
二、中 枢 抑 制(central inhibition)
(一)突触后抑制
如果突触后膜发生超极化,即产生抑制性突触后电位,使突触后神经元兴奋性降低,不易去极化而呈现抑制。这种抑制就称为突触后抑制(postsynaptic inhibition)。突触后抑制根据神经元联系的方式不同,又可分为传入侧支性抑制和回返性抑制。
1.传入侧支性抑制(collateral inhibition)是指一条感觉传入纤维的冲动进入脊髓后,一方面直接兴奋某一中枢神经元,另一方面通过其侧支兴奋另一抑制性中间神经元,然后通过抑制性中间神经元的活动转而抑制另一中枢神经元。
2.回返性抑制是指某一中枢的神经元兴奋时,其传出冲动在沿轴突外传的同时,又经其轴突侧支兴奋另一抑制性中间神经元,后者兴奋沿其轴突返回来作用于原先发放冲动的神经元。回返性抑制(recurrent inhibition)的结构基础是神经元之间的环式联系,其典型代表是脊髓内的闰绍细胞对运动神经元的反馈抑制,脊髓腹角运动神经元在发出轴突支配骨骼肌时,其轴突在尚未离开脊髓腹角灰质前发出侧支支配腹角灰质中一种小的神经细胞—闰绍细胞。闰绍细胞是个抑制性中间神经元,它兴奋时使原发放冲动的运动神经元发生抑制。
(二)突触前抑制
当突触后膜受到突触前轴突末梢的影响,使后膜上的兴奋性突触后电位减小,导致突触后神经元不易或不能兴奋而呈现抑制,称为突触前抑制(presynaptic inhibition)。
第三节 神经系统的感觉机能
- 感受器
(一)感受器的分类
感受器可分为外感受器(exteroceptor)和内感受器(interceptor)两大类。每大类又可分为几小类,如下所示:

(二)感受器的一般生理特性
1.适宜刺激(adequate stimulus)一般说来,每一种感受器都有它的适宜刺激。如视网膜的适宜刺激为光波,内耳柯蒂氏器的适宜刺激是机械波,皮肤上温度感受器的适宜刺激是温度变化等等。
2.感受器的阈值及其换能作用感受器接受刺激发生兴奋,使刺激的能量转化为神经上的电活动,这就是感受器的换能作用(transduction of receptor)。
3.刺激强度与神经冲动的关系在一定刺激强度范围内,感受器受到刺激时,冲动发放的频率与刺激强度的对数成正比。较弱的阈上刺激,冲动发放频率较低,只能引起少数(感受性较高)感觉神经元兴奋;较强的阈上刺激,冲动发放频率较高,能使更多的感觉神经元(感受性较低)产生兴奋。
4.感受器的适应以恒定的刺激强度持续作用于感受器时,将引起它的传入神经纤维上的冲动频率逐渐降低,这一现象称为感受器的适应。不同感受器的适应速度不同,例如,痛觉感受器和颈动脉窦的压力感受器都是适应很慢的感受器;而嗅觉和触觉感受器的适应却很快。
5.感受器的反馈调节如在视网膜、耳蜗螺旋器、前庭器官的壶腹嵴、肌梭等感受器都证明有传出神经支配。这种传出神经纤维的调节作用,多数是属于抑制性的。它们是通过反馈作用来实现的一种自身调节。
二、脊髓的感觉传导通路
(一)浅感觉传导路
传导皮肤和粘膜的痛觉、温度觉和轻触觉冲动。由三级神经元组成。现分述如下:
躯干、四肢的浅感觉由传入神经传至脊髓背角,在背角灰质区换神经元,再发出纤维在中央管下交叉到对侧,分别经脊髓丘脑侧束(痛、温度觉)和脊髓丘脑腹束(轻触觉)前行达丘脑。再由丘脑更换第三级神经元,投射到大脑皮质的躯体感觉区。此传导路径概括如下图:
头面部的浅感觉经三叉神经传入脑桥后,其中传导轻触觉的纤维止于三叉神经核,而传导痛、温度觉的纤维止于三叉神经脊束核。二者换元后,交叉到对侧前行,组成三叉丘系,经脑干各部行至丘脑更换第三级神经元投射到大脑皮质的躯体感觉区。此路径概括如下图:
(二)深感觉传导路
传导肌、腱、关节等的本体感觉和深部压觉的冲动。由这些部位的感受器所发出的冲动经脊神经传入脊髓背角,沿同侧背索前行抵达延髓的薄束核和楔束核。在此更换神经元并发出纤维交叉到对侧,经内侧丘系达丘脑,在丘脑换第三级神经元投射到大脑皮质的躯体感觉区。此路径概括如下图:
可见,脊髓在传导感觉冲动的途径中,都有一次交叉。浅感觉传导路是先交叉再前行;深感觉传导路是先前行再交叉。因此,在脊髓半断离的情况下,浅感觉的障碍发生在断离的对侧,而深感觉的障碍发生在断离的同侧。
三、丘脑及其感觉投射系统
丘脑是感觉传导的接替站。来自全身各种感觉的传导通路(除嗅觉外),均在丘脑内更换神经元,然后投射到大脑皮质。在丘脑内只对感觉进行粗略的分析与综合,丘脑与下丘脑、纹状体之间有纤维互相联系,三者成为许多复杂的非条件反射的皮质下中枢。
(一)丘脑核团的分类
丘脑的核团大致分3类。
1.第一类(感觉接替核)接受感觉的投射纤维,换元后投射到大脑皮质的感觉区。如后腹核、内侧膝状体、外侧膝状体等。
2.第二类接受由第一类核团和其它皮质下中枢来的纤维(但不直接接受感觉的投射纤维),换元后投射到大脑皮质某一特定区域。如外侧腹核接受小脑、苍白球来的纤维,投射到大脑皮质运动区等。
3.第三类(主要是髓板内核群)是丘脑的古老部分,这类细胞没有直接投射到大脑皮质的纤维,但可间接地通过多突触接替,换元后弥散地投射到整个大脑皮质,如中央中核、束旁核等。
(二)感觉投射系统及其作用
根据丘脑各核团向大脑皮质投射纤维特征的不同,丘脑的感觉投射系统可分为特异性投射系统和非特异性投射系统。
1.特异性投射系统(specific projection system)
概念:从机体各种感受器发出的神经冲动,进入中枢神经系统后,由固定的感觉传导路,集中到达丘脑的一定神经核(嗅觉除外),由此发出纤维投射到大脑皮质的各感觉区,产生特定感觉。这种传导系统叫做特异性投射系统。
作用:丘脑是特异性传导系统的一个重要接替站,它对各种传入冲动进行汇集,并作初步的分析和综合,产生粗略的感觉,但对刺激的性质和强度,则不能进行精确的分析。
2.非特异性投射系统(unspecific projection system)
概念:各种感觉冲动进入脑干网状结构后,经过许多错综复杂交织在一起的N元的彼此相互作用,就失去了各种感觉的特异性,因而投射到大脑皮质就不再产生特定的感觉。
此系统的作用:一是激动大脑皮质的兴奋活动,使机体处于醒觉状态,所以非特异性投射系统又叫脑干网状结构上行激动系统(ascending activating system)。
四、大脑皮质的感觉分析功能
不同的感觉在大脑皮质内有不同的代表区。各感觉区之间在功能上密切联系,协同活动,产生各种复杂的感觉。
1.躯体感觉区定位于大脑皮质顶叶。低等哺乳类如兔、鼠等的躯体感觉区与躯体运动区基本重合在一起,统称感觉运动区(sensorimotor area)。动物愈高等,躯体感觉区与躯体运动区愈明显分离。
躯体感觉在大脑皮质的投影有以下规律:
①具有左右交叉的特点,但头面部的感觉投影是双侧性的。
②前后倒置,即后肢投影在大脑皮质顶部,且转向大脑半球内侧面,而头部投影在底部。
③投影区的大小决定于感觉的灵敏度、机能重要程度和动物特有的生活方式。
2.感觉运动区即躯体运动区,也是肌肉本体感觉投影区,它与外周神经联系是对侧的。
3.视觉区、听觉区、嗅觉和味觉区见第十三章。
4.内脏感觉区全身内脏感觉神经是混在交感神经和副交感神经中进入脊髓、脑干,更换神经元后,通过丘脑和下丘脑而到达大脑皮质的中央后回和边缘叶。
第四节神经系统对躯体运动的调节
- 脊髓对躯体运动的调节
躯体运动最基本的反射中枢位于脊髓。最基本的脊髓反射(spinal reflex)包括两类:牵张反射和屈肌反射。
(一)牵张反射
概念:无论屈肌或伸肌,当其被牵张时,肌肉内的肌梭就受到刺激,感觉冲动传入脊髓后,引起被牵拉的肌肉发生反射性收缩,从而解除被牵拉状态,这叫做牵张反射(stretch reflex)。一般分为腱反射和肌紧张。
1.腱反射(tendon reflex)是指快速牵拉肌腱时发生的牵张反射。例如,敲击股四头肌腱时,股四头肌发生收缩,膝关节伸直,这叫做膝反射(knee jerk)。敲击跟腱时,引起腓肠肌收缩,跗关节伸直,这叫做跟腱反射。
2.肌紧张是指缓慢地持续牵拉肌腱时所发生的牵张反射。牵张反射的发生,是通过脊髓中枢的兴奋性突触和抑制性突触的双重作用完成的。
(1)兴奋性突触的作用位于骨骼肌纤维间的肌梭(muscle spindle),因突然被牵拉所产生的冲动,沿传入神经元进入脊髓,其中一部分纤维和脊髓腹角α运动神经元发生兴奋性突触联系,使α运动神经元产生大量冲动传到肌纤维,使之猛烈收缩,因而整条肌肉缩短。
(2)抑制性突触的作用包括直接(传入侧支性抑制)和间接(回返性抑制)两种抑制方式。
(二)屈肌反射和对侧伸肌反射
以伤害性刺激施与一侧后肢的下部,如针刺激左(或右)侧后肢跖部皮肤时,就可引起该肢屈曲,这种现象叫做屈肌反射。如果刺激很强,除本侧肢体发生屈曲外,同时引起对侧肢体伸直,以支持体重,这种对侧肢体伸直的反射叫做对侧伸肌反射(crossed extensor reflex)。
上述两种反射的生理意义:在于被刺激侧肢体屈曲,以躲避伤害作用,对侧肢体伸直,以维持机体重心不致跌倒。这些都是比较原始的防御性反射(defense reflex)。
二、脑干对肌紧张的调节
脑干包括延髓、脑桥和中脑。脑干有较多的神经核以及与这些核相联系的前行和后行神经传导路,还有纵贯脑干中心的网状结构。脑干网状结构是中枢神经系统中重要的皮质下整合调节机构,有多种重要功能。其中对牵张反射和姿势反射等躯体运动就有着重要的整合调节作用。
(一)脑干网状结构对牵张反射的调节
脑干网状结构对脊髓运动神经元具有易化和抑制两方面的作用,尤其是对伸肌的牵张反射的调节作用。
1.脑干网状结构后行系统的机能 延髓网状结构的腹内侧部兴奋时,发放冲动到脊髓,能抑制四肢伸肌的牵张反射。如以电流刺激这一部位,原来进行中的腿部伸直动作即被制止,四肢肌肉紧张性立即下降,因此,这部分结构及其后行神经路径称为脑干网状结构后行抑制系统。
在正常机体内,脑干对脊髓躯体运动神经元的后行作用,包括易化和抑制两方面,而且经常保持着动态平衡,从而使全身骨骼肌的紧张性收缩保持适当的强度,躯体运动也得以正常进行,当病变造成这两个系统之间的关系失调时,将出现肌紧张亢进或减弱。
2.去大脑僵直(decerebrate rigidity)将动物麻醉并暴露脑干,在中脑前、后丘之间切断,造成所谓去大脑动物,使脊髓仅与延髓、脑桥相联系,动物则出现全身肌紧张(特别是伸肌)明显加强。表现为四肢僵直,头向后仰,尾巴翘起,躯体呈角弓反张状态。这种现象叫做去大脑僵直。
去大脑僵直的的机制是:一方面,网状结构的后行抑制系统由于失去了大脑皮质和尾状核后行抑制性冲动的控制,其抑制作用相对地减弱;另一方面,网状结构的易化系统和前庭核的活动又有所加强;两方面效应相结合,四肢伸肌及所有抗重力肌肉群的牵张反射便处于绝对的优势。
3.两种去大脑僵直
α僵直:由于高位中枢的后行性作用,直接或间接通过脊髓中间神经元提高α运动神经元的活动,从而导致肌紧张加强而出现僵直,这称为α僵直。
γ僵直:由于高位中枢的后行性作用,首先提高脊髓γ运动神经元的活动,使肌梭的敏感性提高而传入冲动增多,转而使脊髓α运动神经元的活动提高,从而导致肌紧张加强而出现僵直,这称为γ僵直。
(二)脑干对姿势反射的调节
1.状态反射当动物头部在空间的位置改变或头部与躯干的相对位置改变时,反射性地改变躯体肌肉的紧张性,从而形成各种形式的状态,叫做状态反射(attitudinal reflex)。
2.翻正反射动物摔倒时,自行翻转起立,恢复正常站立姿势,叫做翻正反射(righting reflex)。这种反射比状态反射复杂。如将猫四脚朝天,从空中坠下时,首先是头颈扭转,然后前肢和躯干紧接着也扭转过来,最后后肢也扭转过来,当坠到地面时,先由四肢着地。
三、大脑皮质对躯体运动的调节
(一)大脑皮质运动区
大脑皮质的某些区域与骨骼肌运动有着密切关系。运动区对骨骼肌运动的支配有如下特点:
①一侧皮质支配对侧躯体的骨骼肌,两侧呈交叉支配的关系,但对头面部肌肉的支配大部分是双侧性的。
②具有精细的功能定位,即对一定部位皮质的刺激,引起一定肌肉的收缩。而这种功能定位的安排,总的呈倒置的支配关系。即支配后肢肌肉的定位区靠近中央,支配前肢和头部肌肉的定位区在外侧。
③支配不同部位肌肉的运动区,可占有大小不同的定位区,运动较精细而复杂的肌群(如头部),占有较广泛的定位区,而运动较简单而粗糙的肌群(如躯干、四肢)只有较小的定位区。
(二)锥体系统
锥体系统(pyramidal system)是指由大脑皮质发出并经延髓锥体而后行达脊髓的传导束。
锥体系统是大脑皮质后行控制躯体运动的直接通路。调节单个肌肉的精细动作。
(三)锥体外系统
皮质下某些核团(如尾核、壳核、苍白球、黑质、红核等)有后行通路控制脊髓运动神经元的活动。其通路在延髓锥体之外,故叫锥体外系统(extrapyramidal system)。锥体外系统的机能主要是协调全身各肌肉群的运动,保持正常姿势。
- 神经系统对内脏活动的调节
一、交感和副交感神经的特征
支配内脏器官的交感神经和副交感神经与支配骨骼肌的躯体神经相比,具有以下结构和生理上的特征。
1.交感神经起自脊髓胸腰段侧角,经相应的腹根传出,通过白交通支进入交感神经节,副交感神经的起源比较分散,其中一部分起自脑干有关的副交感神经核(动眼神经中的副交感神经纤维起自中脑缩瞳核;面神经和舌咽神经中的副交感神经纤维分别起自延髓上唾液核和下唾液核;迷走神经中的副交感神经纤维起自延髓迷走背核和疑核),另一部分起自脊髓荐部,相当于侧角的部位。
2.植物性神经的纤维离开中枢神经系统后,不直接到达所支配的器官,先终止于神经节并换神经元,再发出轴突到达器官。因此,中枢的兴奋,通过植物性神经传到效应器,必须经过两个神经元,即由中枢发出到神经节的纤维,叫做节前纤维(preganglionic fibers);由神经节到效应器的纤维,叫做节后纤维(postganglionic fibers)。交感神经节离开效应器较远。其节前纤维短,节后纤维长。
3.当刺激交感神经节前纤维时,效应器发生反应的潜伏期长。刺激停止后,它的作用可持续几秒或几分钟,刺激副交感神经节前纤维引起效应器活动时,其潜伏期短。刺激停止后,作用持续时间也短。
二、交感和副交感神经的功能
如下表:
9-4植物性神经的生理作用
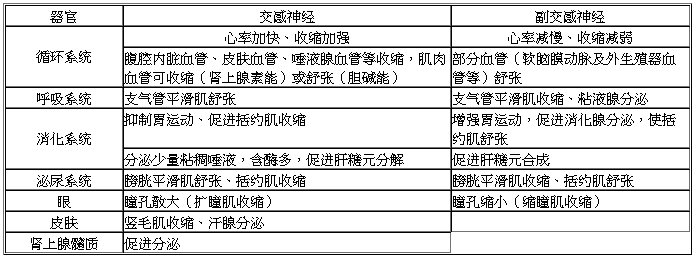
1、在具有双重神经支配的器官中,它们对同一器官的作用,往往具有相互颉颃的性质。
例如,对于心脏,迷走神经具有抑制作用,而交感神经则具有兴奋作用;对胃肠活动,迷走神经具有兴奋作用,而交感神经则具有抑制作用。这两种神经从正、反两方面调节器官的活动,使器官的活动水平能适应机体的需要。
2、植物性神经对器官的支配,一般具有持久的紧张性作用。
例如,切断支配心脏的迷走神经时,心率就加快。这表明迷走神经经常有紧张性冲动传出来,对心脏发生持续的抑制作用;又如切断心交感神经时,则心率减慢,这表明心交感神经的活动也具有紧张性。
3、植物性神经的外周性作用与效应器本身的机能状态有关。
例如,胃肠如果原来处于收缩状态,则刺激迷走神经可引起舒张,如原来处于舒张状态,则刺激迷走神经却引起收缩;又如刺激交感神经可导致动物无孕子宫的运动受到抑制,而对有孕子宫则可加强运动。这些说明植物性神经的作用随着支配器官本身的机能状态,可以互相转化。
4、交感神经系统(sympathetic nervous system)的活动,一般较广泛往往不是波及个别神经纤维及其所支配的效应器,而常以整个系统来参与反应。例如,在动物剧烈运动、窒息、失血或寒冷等情况下,由于反射地兴奋交感神经系统,机体出现心率加快、收缩加强,皮肤和腹腔内脏血管收缩、增加心输出量、血压升高、血液循环加快;支气管舒张、增加通气量;肾上腺素分泌增加,肝糖原分解加速,血糖升高等等。
5、由于交感神经系统活动加强时常伴随肾上腺素分泌增多,因此,往往将这一活动系统叫做“交感—肾上腺”系统。又由于迷走神经活动加强时,常伴随胰岛素分泌增加,因此又将这一活动系统叫做“迷走—胰岛素”系统。在应激情况下,不但交感—肾上腺系统发生广泛兴奋,迷走—胰岛素系统也广泛兴奋,但前者较后者作用强。所以,后者效应被掩盖,而不易察觉。
三、内脏活动的中枢性调节
(一)脊髓
交感神经和部分副交感神经,起源于脊髓灰质的侧角内,因此,脊髓是调节内脏活动的最基本中枢,通过它可以完成简单的内脏反射活动。例如排粪、排尿、血管舒缩以及出汗和竖毛等活动。但是这种反射调节功能是初级的,不能更好地适应生理机能的需要,在正常时脊髓受高级中枢的调制(modulation)。
(二)脑干
部分副交感神经由脑干发出,支配头部的腺体、心脏、支气管、食管、胃肠道等。同时在延髓中还有许多重要的调节内脏活动的基本中枢。如调节呼吸运动的呼吸中枢;调节心血管活动的心血管运动中枢;调节消化管运动和消化腺活动的中枢等。可完成比较复杂的内脏反射活动。延髓一旦受到损伤,可导致各种生理活动失调,严重时可引起呼吸或心搏停止,因此延髓被称为“生命中枢”所在地。
(三)下丘脑
下丘脑是大脑皮质下调节内脏活动的较高级中枢,它能够进行细微和复杂的整合作用,使内脏活动和其它生理活动相联系,以调节体温、水平衡、摄食等主要生理过程。
1.体温调节下丘脑是体温调节的主要中枢所在地。当体内、外温度发生变化时,可通过体温中枢对产热或散热机能进行调节,使体温恢复正常和经常保持相对稳定状态。
2.水平衡调节下丘脑的视上核和室旁核是水平衡调节中枢。它调节水平衡,包括两方面。一是控制抗利尿激素的合成和分泌,另一是控制饮水。如血浆渗透压异常升高时,可引起垂体后叶释放抗利尿激素进入血液,随血液循环到达肾脏,促进远曲小管和集合管对水分的重吸收,同时产生渴感,驱使动物大量饮水,共同调节水平衡。
3.摄食行为调节下丘脑存在有摄食中枢(feeding center)和饱中枢(satiety center)。如果破坏摄食中枢,动物拒绝摄食;破坏饱中枢,动物食欲大增,逐渐肥胖。实验证明,血糖水平的高低可能调节摄食中枢和饱中枢的活动,这主要取决定于神经元对葡萄糖的利用程度。
4.内分泌腺活动的调节下丘脑有许多神经元具有分泌机能,可分泌多种激素,进入血液,并通过垂体门脉循环到腺垂体,促进或抑制腺垂体各种激素的合成和分泌,进而调节其它内分泌腺的活动。
(四)大脑边缘系统
大脑半球内侧面皮质与脑干连接部和胼胝体旁的环周结构,叫做“边缘叶”。它与大脑半球外侧面皮质相比,这些结构属于进化上比较古老的皮质,故又叫旧皮质,边缘叶包括扣带回、胼胝体回、海马沟与海马回等。
由于边缘叶在结构上和大脑皮质的岛叶、颞极、眶回等,以及杏仁核、隔区、下丘脑、丘脑前核等密切相关。于是人们常把边缘叶连同这些结构统称为边缘系统(limbic system)。
大脑边缘系统是内脏活动的重要调节中枢,而且还与情绪(emotion)、记忆功能有关。
第六节脑的高级神经活动
一般把条件反射叫做高级神经活动(higher nervous activity)。
一、条 件 反 射
(一)非条件反射(unconditioned reflex)和条件反射(conditioned reflex)
非条件反射是动物在种族进化过程中,适应变化的内外环境通过遗传而获得的先天性反射,是动物生下来就有的。这种反射有固定的反射途径。反射比较恒定,不易受外界环境影响而发生改变,只要有一定强度的相应刺激,就会出现规律性的特定反应,其反射中枢大多数在皮质下部位。
例如,饲料进入动物口腔,就会引起唾液分泌;机械刺激角膜就会引起眨眼等都属于非条件反射。
条件反射是动物在出生后的生活过程中,适应于个体所处的生活环境而逐渐建立起来的反射,它没有固定的反射途径,容易受环境影响而发生改变或消失。因此,在一定的条件下,条件反射可以建立,也可以消失,
(二)条件反射的形成
条件反射是建立在非条件反射基础上的,形成条件反射的基本条件为:
第一,无关刺激与非条件刺激在时间上的反复多次结合。这个结合过程叫做强化(reinforcement)。
第二,无关刺激必须出现在非条件刺激之前或同时。
第三,条件刺激的生理程度比非条件刺激要弱。例如动物饥饿时,由于饥饿加强了摄食中枢的兴奋性,食物刺激的生理强度就大大提高,从而容易形成条件食物反射。
(四)条件反射的消退
已形成的条件反射,如果在给予条件刺激时,再不伴用非条件刺激强化,久而久之,原来的条件反射逐渐减弱,甚至不再出现,这称为条件反射的消退。
(五)条件反射的生理学意义
条件反射的建立,极大地扩大了机体的反射活动范围,增加了动物活动的预见性和灵活性,从而对环境变化更能进行精确的适应。
二、神经活动的类型
(一)家畜的基本神经型
1.兴奋型其特点是兴奋和抑制都很强,兴奋更占优势。这类动物的表现是急躁、暴烈、不受约束和带有攻击性,它们能迅速地建立比较巩固的条件反射,但条件反射的精细度很差,即对类似刺激辨别能力很弱。
2.活泼型其特点是兴奋和抑制都强,且均衡发展,互相转化比较容易且迅速。这类动物表现为活泼好动,对周围发生的微小变化能迅速发生反应。它们形成条件反射很快,能精细的辨别相似的刺激,能适应环境的复杂变化,是生理上最好的神经型。
3.安静型其特点是兴奋和抑制都强,发展也比较平衡,但互相转化比较困难而缓慢。这类动物表现为安静、细致、温驯和有节制,对周围变化反应冷淡。它们能很好的建立精细的条件反射,但形成的速度较慢。
4.抑制型其特点是兴奋和抑制都很弱。一般更容易表现抑制。这类动物胆怯而不好动,易于疲劳,常常畏缩不前和带有防御性,它们一般不易形成条件反射,形成后也不巩固。它们不能适应变化复杂的环境,也难于胜任比较强和持久的活动。
(二)神经型的形成
家畜的神经型既取决于神经系统的遗传特性,又取决于个体后天的生存条件。神经系统的遗传特性实际上就是形成条件反射的可能性,它包括形成的速度、强度和稳定性等方面的个体差异。至于真正形成条件反射或形成什么样的条件反射,必须要在后天的环境影响下才能完成,所以家畜的神经型是皮质功能的遗传性与同围环境影响相结合的产物。
(三)神经型的实践意义
以马和耕牛来说,活泼型的使役能力高、挽力大、速度快,能迅速适应使役条件的变化,稍驱赶就加快运动;安静型的使役能力好,挽力也大,但动作缓慢,常常需要驱赶;兴奋型在使役强度不大时表现良好,使役强度增大时,则表现能力不定,对驱赶表现反抗;抑制型的使役能力很差,挽力小、速度慢、不耐久,对驱赶反应迟纯。
以猪来说,安静型的个体容易肥育,而兴奋型的个体则难以肥育。
第四章 血液循环
·循环系统由心脏、血管组成的密闭管道系统。血液在其中按一定方向流动,称为血液循环。血液循环的主要功能是完成体内的物质运输,运输代谢原料和代谢产物,使机体新陈代谢能不断进行;体内各内分泌腺分泌的激素,或其它体液性因素,通过血液的运输,作用于相应的靶细胞,实现机体的体液调节;机体内环境理化特性相对稳定的维持和血液防卫机能的实现,也都有赖于血液的不断循环流动。
·血液先由左心室泵出,经动脉至毛细血管,在此与组织细胞进行物质交换,即送去养分、氧气,并带走代谢产物,然后经静脉回流入右心房。上述流动过程叫体循环(systemic circulation),因其循环线路长,故也称之为大循环。
·血液经右心房流至右心室,再由右心室射出,经肺动脉及肺毛细血管,并在此与肺泡内气体进行气体交换,即放出二氧化碳,吸取氧气。然后,含氧丰富的血液经肺静脉回流至左心房,进入左心室,这个流动过程,叫做肺循环(pulmonary circulation)。因其循环线路短,故也称之为小循环。
·除上述两个循环路线外,部分组织液进入另一套封闭的管道系统,形成淋巴液。后者经小淋巴管、大淋巴管,再经左侧胸导管和右颈淋巴干分别进入左、右锁骨下静脉,形成淋巴回流,淋巴回流可视为体循环的一个旁支。
第一节心脏的生理活动
一、心肌细胞的生物电现象
心肌细胞在细胞膜两侧存在着电位差,这种电位差称为跨膜电位,它包括静息状态下的静息电位和兴奋时的动作电位。
(一)心室肌细胞的跨膜电位及其产生原理
1.静息电位当完整无损的心室肌处于静息状态时,其表面各点的电位相等,唯膜内外存在跨膜电位差,用微电极可检测到其静息电位约为-90mV,并表现膜外带正电,膜内带负电的极化状态。心室肌细胞的静息电位及其形成原理,也是由于K+向细胞膜外流动所产生的K+跨膜电位或平衡电位。
2.动作电位是指心室肌细胞受到刺激而发生兴奋时,其静息电位经历去极化与复极化的转变过程。
主要经历以下几个时期:
(1)去极过程(0期)为动作电位的上升支。持续的时间很短,仅1~2ms。但其去极化的速度很快,
0期主要由于Na+的快速内流所致。
(2)复极过程包括四个时期:
①快速复极初期(1期)在复极初期,仅出现部分复极,膜内电位由+30mV左右迅速下降到0mV左右,故称为快速复极初期,占时约10ms。0期除极和1期复极这两个时期的膜电位变化都很快,记录图形上表现为尖峰状,故称为锋电位。研究表明,K+是该外向电流的主要离子成分。
②平台期(2期) 当1期复极结束后,膜内电位达到0mV左右, 复极过程变得非常缓慢,膜内电位基本上停滞于0 mV左右,细胞两侧呈等电位状态,记录图形比较平坦,故复极2期又称为坪或平台期,持续约100~150ms,是整个动作电位持续时间长的主要原因,是心室肌细胞以及其它心肌细胞的动作电位区别于骨骼肌和神经纤维的主要特征。通常认为,平台期是由Ca2+(以及Na+)的内向离子流而形成的。
③快速复极末期(3期)平台期后,由于Ca2+通道完全失活,内向离子流终止,外向K+流进一步增强,因而膜的复极加速,导致膜电位快速复极化直至完成复极。此期主要是快钾通道(在平台期已被激活)开放,K+依其膜内外的浓度差快速外流的结果。此期约占时100~150ms。
④静息期(4期) 3期之后膜电位稳定在-90mV的水平。此时的膜电位虽已恢复到静息水平,但离子分布状态尚未复原。心肌细胞膜上存在有Na+—K+泵,通过泵的作用,将Na+的外运与K+的内运相耦联起来, 形成Na+—K+交换,实现Na+、K+的主动转运。关于Ca2+的逆浓度梯度外运,有人认为与Na+顺浓度梯度的内流相耦联,形成Na+-Ca2+交换。
由于离子主动转运过程的电荷量基本相等,对心室肌细胞而言,4期膜电位基本保持稳定的静息水平,故称静息电位,四期又称为静息期。
(二)窦房结细胞跨膜电位的特点
窦房结细胞是一种自律细胞(autorhythmic cell),它在没有外来刺激的情况下,也会自动去极化。因此,其跨膜电位的变化,尤其在动作电位的产生过程中,有别于其它心肌细胞。窦房结细胞的动作电位只表现为0、3、4三个时期。
(三)心肌的快反应细胞和慢反应细胞
1.快反应细胞此类细胞有心房肌细胞、心室肌细胞、优势传导通路、房室束和浦肯野氏纤维。快反应细胞的0期去极化由Na+的快速内流引起。其特点是:动作电位的幅度较大,上升的速度较快,传播的速度也较快。其中心房肌和心室肌细胞的4期稳定,不能发生自动去极化,没有自律性,因此又叫快反应非自律性细胞。
2.慢反应细胞此类细胞包括窦房结、房室交界的房结区和结希区等的细胞。慢反应细胞的0期去极化由Ca2+内流所引起,其电位幅度较小,0期去极化速度和动作电位传播速度较慢。慢反应细胞的4期不稳定,能发生自动去极化,因此具有自律性。
二、心肌的生理特性
(一)自动节律性
组织细胞在没有外来刺激的条件下,能自动地发生节律性兴奋的特性,称为自动节律性,简称自律性(autorhythmicity)。具有自动节律性的组织或细胞,叫自律组织或自律细胞。
窦房结的自律细胞,依组织学的特点定名为苍白细胞(pale cell),简称P细胞。4期自动除极是自律性的基础,窦房结P细胞的自律性最高,心脏始终是依照当时情况下自律性最高的部位所发出的兴奋来进行活动的。
概念:
- 正常起搏点:窦房结是主导整个心脏兴奋和跳动的正常部位,称之为正常起搏点。
- 潜在起搏点:其他部位的自律组织并不表现出它们自身的自动节律性,只是起着兴奋传导的作用,称之为潜在起搏点。
3、异位起搏点:在某种异常情况下,窦房结以外的自律组织也可以自动产生兴奋,而心房或心室则依从当时情况下节律性最高部位的兴奋而跳动,这些异常的起搏部位称为异位起搏点。
4、窦性心律(sinus rhythm):正常心搏节律是由自律性最高处窦房结发出冲动引起,故称窦性心律(sinus rhythm)。
5、异位心律(ectopic rhythm),指由窦房结以外的自律细胞取代窦房结而主宰心搏节律。
决定和影响自律性的因素:
1.最大复极电位与阈电位之间的差距
2.4期自动除极速度
(二)兴奋性
所有心肌细胞都具有兴奋性。
决定和影响兴奋性的因素:
(1) 静息电位水平静息电位(自律细胞为最大复极电位)绝对值增大时,距离阈电位的差距就加大,引起兴奋所需的刺激阈值增大,表现为兴奋性降低。反之,静息电位绝对值减小时,距阈电位的差距缩小,所需的刺激阈值减小,兴奋性增高。
(2) 阈电位水平阈电位水平上移,则与静息电位之间的差距增大,引起兴奋所需的刺激阈值增大,兴奋性降低。反之亦然。
(3) Na+通道的性状
心室肌细胞一次兴奋过程中,其兴奋性的变化可分以下几个时期:
(1) 绝对不应期与有效不应期心肌细胞发生一次兴奋后,由动作电位的去极相开始到复极3期膜内电位达到约-55mV这一段时期内,如果再受到第二个刺激,则不论刺激强度多大,都不发生反应,称为绝对不应期。
(2)相对不应期指膜电位从-60 mV继续复极化至-80 mV期间,此时Na+通道的开放能力已逐渐恢复,但仍然低于正常,故只对阈上刺激能产生动作电位,而且表现为0期幅度和速度均较正常为小,传导速度也较慢。
(3) 超常期心肌细胞继续复极,膜电位从-80mV继续恢复至-90mV的极短时间内,Na+通道已恢复到可以再激活的状态,亦即兴奋性已基本恢复,而且此时的膜电位比正常电位更接近阈电位,因此以稍低于阈刺激的阈下刺激就足以使心肌兴奋,表明此期的兴奋性超过正常,故名超常期。
(三) 传导性
传导性(conductivity)是指心肌细胞兴奋产生的动作电位能够沿着细胞膜传播的特性。心肌细胞的兴奋可以通过细胞间的闰盘(缝隙连接),从一个细胞扩布到其相邻的细胞,使心房、心室各为功能上的合胞体而表现为左、右心房或心室的同步兴奋和收缩。
心脏内兴奋传导的途径是:窦房结产生的兴奋,经过渡细胞传至心房,通过优势传导通路传导到房室交界(房结区、结区、结希区),再经房室束、房室束支、浦肯野氏纤维网至心室肌。
心肌的传导性主要受0期除极的速度、幅度、静息电位水平和阈电位水平的影响。
(1) O期除极的速度和幅度0期除极速度快、幅度高,则邻近安静部位爆发兴奋所需的时间就短,兴奋传导就快;反之兴奋传导就慢。
(2) 静息电位或最大舒张电位的水平在一定范围内,静息时的跨膜电位梯度(即膜内外的电位差)是推动Na+内流和0期去极化的重要因素。静息电位绝对值大,跨膜电位梯度大,就促进Na+内流,加快0期去极化的速度,因而兴奋传导快, 传导性升高;静息电位减小则相反,兴奋传导减慢,传导性降低。
(3) 阈电位水平阈电位水平下移(绝对值大),则从静息电位到达阈电位的差距减小,扩布性兴奋产生的时间缩短,故兴奋传导速度加快;反之,阈电位水平上移(绝对值小),则兴奋传导慢。
(四)收缩性
心肌细胞的收缩性(contractility)有其自已的特点。
1.对细胞外液中Ca2+浓度的依赖性Ca2+是一切肌细胞兴奋-收缩耦联的媒介。肌收缩所需的Ca2+,除从终末池释放外,尚需细胞外液中的Ca2+通过肌膜和横管内流(心室肌动作电位2期的Ca2+内流)获得。兴奋之后,肌浆中的Ca2+一部分返回终末池储存,另一部分转运出细胞。心肌细胞的横管系统比骨骼肌的发达,这对Ca2+的内流提供了更大的面积。在一定范围内,细胞外液中的Ca2+浓度升高,兴奋时内流的Ca2+量增多,心肌收缩增强;反之,细胞外液中的Ca2+浓度降低。则心肌收缩减弱。当细胞外液中Ca2+浓度降得很低,甚至无Ca2+时,心肌肌膜虽仍能兴奋产生动作电位,但细胞却不能收缩,这一现象,称为“兴奋—收缩脱耦联”。
2.同步收缩(全或无收缩)心房和心室内特殊传导组织的传导速度快,而心肌细胞之间的闰盘(缝隙连接)又为低电阻区, 因此,兴奋在心房和心室内传导速度快,兴奋几乎同时到达所有的心房肌细胞或心室肌细胞,从而引起整个心房或心室同时收缩(同步收缩)。同步收缩的力量大,有利于射血。由于同步收缩,所以心房或心室要么不收缩,要么整个心房或整个心室一起收缩,这种收缩现象称为“全或无收缩”。
3.不发生强直收缩心肌兴奋性周期变化的特点是有效不应期特别长,相当于整个收缩期加舒张早期。在此期间,任何强刺激都不能引起心肌收缩。所以每次收缩后必有舒张,始终保持着收缩与舒张交替的节律活动。
4.期前收缩与代偿性间歇
在有效不应期之后,若给予一次实验条件下的人工刺激,或在病理情况下有来自异位节律点的刺激,则可引起心室肌收缩。由于这种收缩发生在窦房结兴奋所引起的正常收缩之前,故称为期前收缩或额外收缩(compensatory pause),也称早搏(premature pacemaker)。
由于期前收缩的出现,使紧接而来的窦房结兴奋往往落在期前收缩的有效不应期内, 以致心室不能表现收缩反应,必须等到下一次窦房结的兴奋传来时,心室才发生收缩。这样,在一次期前收缩之后,常有一段较长的心脏舒张期,称为代偿性间歇(compensatory pause)。
三、心动周期及其中的各种变化
(一) 心动周期与心率
1.心动周期心动周期是指心脏每收缩、舒张一次所占的时间。
2.心率心率(heart rate)为心搏频率的简称,以每分钟心搏次数(次/min)为单位。
(二)心脏的射血过程及心内的压力和容积变化
1.心房收缩期
2.心室收缩期包括等容收缩相以及快速和减慢射血相。
(1)等容收缩相
(2)射血相
3.心室舒张期包括等容舒张相和心室充盈相,后者又再细分为快速充盈、减慢充盈和心房收缩充盈三个时相。
(1) 等容舒张相
(2) 心室充盈相
(三) 心输出量(cardiac output)
1.心输出量的概念
通常是指左心室射入主动脉的血量。
心输出量是评价循环系统效率高低的重要指标。一侧心室在每次收缩时射入动脉的血量,叫做每搏输出量(stroke volume)。通常把每搏输出量占心室末期的容积百分比称为射血分数(ejection fraction)。平时所指的心输出量,都是指每分输出量, 通常用每搏输出量和心率的乘积来计算即:心输出量=每搏输出×心率。
2.影响心输出量的因素
(1) 心室舒张末期容量(end diastolic volume)即心射血前容量,它决定于静脉的回流量。
(2)心肌后负荷心肌后负荷是指心室肌收缩时承受的负荷,也即心室收缩面临动脉压的阻力大小。
(3)心率
(四)心音
心音(heart sound)是心脏泵血过程中,由于心肌收缩、瓣膜启闭、血流加速和减速对心血管壁的加压和减压作用,以及形成的涡流等因素引起的机械振动而产生的声响。
在一个心动周期中一般可听到“通-塔”这两个心音,偶尔还可听到第三个心音。
(1) 第一心音发生在心缩期,音调较低,持续时间长,属浊音,在心尖搏动处听得最清楚。
(2) 第二心音发生于心舒期,音调较高,持续时间较短。
(3) 第三心音发生于第二心音之后,是一种低频、低振幅的心音。
(4) 第四心音是与心房收缩有关的一组心室收缩前的振动,故也称心房音。
(五) 心电图
心电图(electrocardiogram)是心电活动由体表描记所得的电位变化曲线,反映心脏兴奋起源以及兴奋扩布于心房、心室的过程。
P波 反映左右心房去极化过程,其波形往往小而圆钝。
QRS波群典型的QRS波群往往包括了3个相连的波:第一个是向下的Q波;第二个是高而尖峭的向上的R波;第三个是一个向下的S波, 它所反映的是左右心室兴奋传播过程的电位变化。
T波是继QRS波群之后的一个波幅较低而持续时间较长的波,它反映心室兴奋后的复极化过程。
U波在T波之后有时出现的一个小波。其产生原因还不太清楚。有人认为是心肌舒张时各部先后产生的负后电位形成的;有人认为是浦肯野氏纤维复极化时形成的。
第二节血管的生理活动
血管可分为动脉、毛细血管和静脉3大类。
1.弹性贮器血管指主动脉、肺动脉主干及其发出的最大的分支。
2.分配血管弹性贮器血管以后到分支为小动脉前的动脉管道,其功能是将血液输送至各组织器官,故称为分配血管。
3.毛细血管前阻力血管小动脉和微动脉的管径小,对血流的阻力大,称为毛细血管前阻力血管。
4.毛细血管前括约肌在真毛细血管的起始部常有平滑肌环绕,称为毛细血管前括约肌(precapillary sphincter)。
5.交换血管(exchange vessels)指真毛细血管,其管壁仅由单层内皮细胞构成,外面有一薄层基膜,故通透性很高,是血管内血液和血管外组织液进行物质交换的场所。
6.毛细血管后阻力血管指微静脉,微静脉因管径小,对血流也产生一定的阻力。
7.容量血管静脉在血管系统中起着血液贮存库的作用,故生理学中常将静脉称为容量血管(capacitance vessels)。
8.短路血管指一些血管床中小动脉和小静脉之间的直接联系。
综上所述,血管系统在体内起着输送、分配、贮存血液、调节血流的功能,参与实现机体与环境之间的物质交换过程。
一、血压的概念
血压(blood pressure)是指血管内血流对于单位面积血管壁的侧压力,即压强。导致血压形成的主要因素有:
- 血液充盈血管
②心脏射血
二、动脉血压与动脉脉搏
(一)动脉血压及其影响因素
动脉血压(arterial blood pressure)一般所说的血压系指体循环的动脉血压,它可决定其它部位血管的血压高低。不同种属动物的动脉血压是不一样的。同种动物,其血压也随年龄、性别及生理状况的不同而不同。
影响动脉血压的因素如下:
(1) 心脏每搏输出量如果每搏输出量增大,心缩期射入主动脉的血量增多,心缩期中主动脉和大动脉内的血量增加,管壁所受的张力增大,故收缩期动脉血压的升高更加明显。
(2) 心率
(3) 外周阻力
(4) 主动脉和大动脉的弹性贮器作用如前所述,由于主动脉和大动脉的弹性贮器作用,动脉血压的波动幅度明显小于心室内压的波动幅度。
(5) 循环血量和血管系统容量的比例
(二) 动脉脉搏
在每个心动周期中,动脉内的压力发生周期性的波动。这种周期性的压力变化可引起动脉血管发生搏动,称为动脉脉搏。
三、静脉血压、静脉血流及静脉脉搏
(一)静脉血压
静脉血压(venous pressure)是指静脉内血液对管壁产生的侧压力。
影响静脉血压的因素非常复杂,主要有以下几项。
(1) 左心室的收缩力
(2) 微动脉的血流量
(3) 右心室的收缩和舒张
(4) 静脉系统的阻力
(5) 外力
(二) 静脉脉搏
右心房缩舒活动时产生的压力变化,可逆向传到近心脏的大静脉,从而出现静脉搏动,称静脉脉搏。
第三节微循环、组织液与淋巴液
一、微循环与毛细血管压
微循环是指微动脉(arteriole)和微静脉 (venule) 之间的血液循环。
(一) 微循环的组成与机能
血液从微动脉流向微静脉可通过3条途径,即直捷通路、迂回通路、动—静脉短路。
1.直捷通路血液从微动脉经后微动脉、通血毛细血管而后回到微静脉,这一条通路叫做直捷通路(thoroughfare channel)。
2.迂回通路血液从微动脉经后微动脉、毛细血管网到微静脉的这条通路叫做迂回通路。
3.动—静脉短路血液从微动脉经动-静脉吻合支(arteriovenous anastomoses)直接回流到微静脉,这一通路叫做动-静脉短路。
二、组织液的生成及其影响因素
(一)组织液的生成
血液在流经微循环营养通路时,在毛细血管动脉端有组织液生成,而在静脉端发生组织液回流,从而实现血液与组织液的物质交换。
成因:毛细血管壁的通透性是前提。动力是毛细血管内外存在的四种力量决定的有效率过压,四种力量是:毛细血管血压、组织液静压、血浆胶体渗透压、组织液胶体渗透压。如图:
(二)影响组织液生成的因素
1.毛细血管血压毛细血管血压升高,可促进组织液的生成
2.血浆胶体渗透压血浆蛋白质含量减少,血浆胶体渗透压降低,有效滤过压增大,导致组织液生成增多,引起组织水肿。
3.毛细血管壁的通透性在过敏反应中,由于局部组织释放大量组胺,使毛细血管壁的通透性加大,部分血浆蛋白渗出,血浆胶体渗透压降低,而组织液胶体渗透压升高,组织液生成增多,出现局部水肿。
4.淋巴回流毛细血管动脉端滤出的液体,一部分通过淋巴回流入血液,若淋巴回流受阻,则组织液积聚起来,出现水肿。
三、淋巴的生成和回流
(一)淋巴液的生成
淋巴液来自组织液。血浆滤过生成的组织液仅90%被重吸收,其余进入毛细淋巴管形成淋巴液。
(二) 淋巴回流
毛细淋巴管内的淋巴液流入集合淋巴管和淋巴结,最后经淋巴导管(胸导管和右淋巴管)进入前腔静脉,加入血液循环。所以淋巴回流系统是组织液向血液循环回流的一个重要辅助系统。
组织内的毛细淋巴管起始于盲端。管腔较大而不规则,管壁为单层内皮细胞,其外无基膜, 相邻内皮细胞边缘有似瓦片状的重叠覆盖,并可向管腔内飘动,形成向管腔内开放的单向瓣膜。组织液及悬浮于其中的微粒,包括激素、酶、抗体以及侵入的细菌和消化吸收的脂肪微滴,都可通过这种瓣膜进入毛细淋巴管,但不能倒流。
淋巴液回流的生理功能,主要是将组织液中的蛋白质分子带回至血液中,并且能清除组织液中不能被毛细血管重吸收的较大的分子以及组织中的红细胞和细菌等。
第四节心血管活动的调节
一、心血管活动的神经调节
(一) 心脏的神经支配
心脏受心交感神经(cardiac sympathetic nerve)和心迷走神经(cardiac vagus nerve)的双重支配。
1.心交感神经及其作用心交感神经的节前神经元位于脊髓第l~5胸段的中间外侧柱内,其轴突末梢释放的递质为乙酰胆碱,它能激活节后神经元细胞膜上的N型胆碱能受体。
2.心迷走神经及其作用支配心脏的副交感神经节前纤维行走于迷走神经干中。这些节前神经元的细胞体位于延髓的迷走神经背核和疑核,在不同的动物中有种间差异。
综上所述,可见心交感神经和心迷走神经对心脏活动的支配效应是相颉颃的。但是,在整体生命活动中,二者的效应既相颉颃又协调统一,具有高度的适应性,神经纤维上的经常性冲动,称为神经紧张。故将支配心脏的心交感神经和心迷走神经的紧张性活动, 分别称为交感紧张(sympathetictone)和迷走紧张(vagul tone)。二者的紧张性常可随着机体生理状态的不同而改变,如动物在相对安静状态下,心迷走紧张占优势,心脏活动减慢减弱;当躯体运动加强时,心交感紧张占主导地位,心脏活动加强加快。以上所发生的适应性变化,主要取决于各级相关中枢之间的高度整合作用。
(二) 调节心血管活动的中枢
1.延髓心血管中枢
一般认为,延髓心血管中枢至少可包括以下四个部位的神经元:①缩血管区引起交感缩血管神经正常的紧张性活动的延髓心血管神经元的细胞体位于延髓头端的腹外侧部,称为C1区。②舒血管区位于延髓尾端腹外侧部Al区(即在Cl区的尾端)的去甲肾上腺素神经元,在兴奋时可抑制C1区神经元的活动,导致交感缩血管紧张降低,血管舒张。③传入神经接替站延髓孤束核的神经元接受由颈动脉窦、主动脉弓和心脏感受器经舌咽神经和迷走神经传入的信息,然后发出纤维至延髓和中枢神经系统其他部位的神经元,继而影响心血管活动。④心抑制区心迷走神经元的细胞体位于延髓的迷走神经背核和疑核。
2.高位心血管中枢在延髓以上的脑干部分以及大脑和小脑中也都存在与心血管活动有关的神经元。它们在心血管活动调节中所起的作用较延髓心血管中枢更加高级,表现为对心血管活动和机体其他功能之间的复杂的整合。
(四) 心血管活动的反射性调节
1.颈动脉窦和主动脉弓压力感受器反射颈动脉窦和主动脉弓血管壁的外膜下,有丰富的感觉神经末梢,主要感受血压变化对血管壁产生的牵张刺激,常称为压力感受器。压力感受器反射是负反馈调节机制。它的生理意义在于使动脉血压保持相对稳定。
2.颈动脉体和主动脉体化学感受器反射颈动脉体和主动脉体可感受血液中的化学变化并发放神经冲动,称之为化学感受器。由该感受器发放神经冲动,所引起的反射活动,称为化学感受器反射(chemoreceptor reflex)。当血液中出现缺氧、CO2分压升高和H+浓度增加时,上述感受器即发放冲动,经窦神经和迷走神经进入延髓,并在孤束核交换神经元,
3.心肺感受器引起的心血管反射心房、心室和肺循环的大血管壁存在着许多感受器,总称为心肺感受器。其中主要有牵张感受器和化学感受器。当心房、心室或肺循环的大血管内压力升高时,或因血容量增大而使心脏或血管壁受到牵张时,心肺感受器可产生兴奋
4.躯体感受器和其它内脏感受器引起的心血管反射刺激躯体传入神经时可以引起各种心血管反射。反射的效应取决于感受器的性质、刺激的强度和频率等。用中低等强度的低频电脉冲刺激骨骼肌传入神经,常可引起降血压效应;而用高强度高频率电刺激皮肤传入神经,则常引起升血压效应。
二、心血管活动的体液调节
心血管活动的体液调节是指血液和组织液中的某些化学物质,对心血管活动所产生的调节作用。按化学物质的作用范围,可分为全身性体液调节和局部性体液调节两大类。
(一) 全身性体液调节
1.肾上腺素和去甲肾上腺素
2.肾素-血管紧张素系统
血管紧张素原(肾素底物,在肝脏合成)
↓ ← 肾素(由近球细胞分泌)
血管紧张素Ⅰ(十肽)
↓←血管紧张素转化酶(主要在肺血管)
血管紧张素Ⅱ(八肽)
↓←血管紧张素酶A
血管紧张素Ⅲ(七肽)
3.血管升压素血管升压素 (vasopressin)是由下丘脑的视上核和室旁核神经元合成,经轴突输送到垂体后叶, 再释放入血。此激素在正常情况下不参与血压调节。只在机体严重失血时,才产生一定的缩血管作用,使血压上升。
(二) 局部性体液调节
1.激肽释放酶-激肽系统激肽释放酶是体内的一类蛋白酶,可使某些蛋白质底物-激肽原分解为激肽。激肽具有舒血管活性,可参与对血压和局部组织血流的调节。
2.组胺组胺是由组氨酸在脱胺酶的作用下生成的。许多组织,特别是皮肤、肺和肠粘膜组织的肥大细胞中,含有大量的组胺。当组织受到损伤或发生炎症以及过敏反应时,均可释放组胺。它的主要作用是使局部毛细血管和微静脉管壁的内皮细胞收缩,彼此分开,使内皮细胞间的裂隙扩大,血管壁的通透性明显增加,导致局部组织水肿。
3.前列腺素各种前列腺素对血管平滑肌的作用是不同的,例如前列腺素E2具有强烈的舒血管作用,前列腺素F2则使静脉收缩。前列环素(即前列腺素I2)是在血管组织中合成的一种前列腺素,有强烈的舒血管作用。
4.阿片肽β内啡肽神经元在大脑基底部和脑干孤束核等均有分布,其轴突投射到其它脑区,所释放的β内啡肽和来自血浆的β内啡肽,作用于某些与心血管活动有关的神经核团,使交感神经活动受到抑制,心迷走神经活动加强,导致血压降低。血浆中的阿片肽还可作用于血管壁上的阿片受体,使血管舒张。
5.心钠素心钠素(cardionatrin)是由心房肌细胞合成和释放的一类多肽。心钠素可使血管舒张,外周阻力降低。也可使每搏输出量减少,心率减慢,故心输出量减少。
第五章 呼吸生理
动物在进行新陈代谢的过程中,需要不断地从外界摄取氧气,氧化营养物质获取能量。同时又必须把在代谢过程中产生的二氧化碳排出体外。机体与外界环境之间的这种气体交换过程称为呼吸(respiration)。
整个呼吸过程由以下三个环节来完成:
①外呼吸,包括肺通气和肺换气。肺通气是指外界气体与肺内气体的交换过程。肺换气是指肺泡气(alveolar gas)与肺泡壁毛细血管内血液间的气体交换过程。
②气体运输,是指机体通过血液循环把肺摄取的氧运送到组织细胞,又把组织细胞产生的二氧化碳运送到肺的过程。
③内呼吸(internal respiration)或组织呼吸,是指血液与组织细胞间的气体交换,它包括组织细胞消耗O2和产生CO2的过程。可见呼吸过程不单靠呼吸系统来完成,还需要血液和血液循环系统的配合。
第一节肺通气
肺通气(pulmonary ventilation)是指肺与外界环境之间的气体交换过程。实现肺通气的器官包括呼吸道、肺泡和胸廓等。呼吸道是沟通肺泡与大气之间的管道;肺泡是肺泡气与血液进行气体交换的主要场所;胸廓的节律性运动是实现肺通气的动力。
一 肺通气的结构基础和功能
(一)呼吸道
呼吸道是气体进出肺的通道,位于胸腔外的鼻、咽、气管称为上呼吸道;位于胸腔内的气管、支气管及其在肺内的分支称为下呼吸道。
(二)肺泡
肺是一对含有丰富弹性组织的气囊,由呼吸性小支气管、肺泡管、肺泡囊和肺泡四个部分组成的功能单位,均具有交换气体的功能,其中以肺泡为主。
1.呼吸膜在电子显微镜下,呼吸膜含有6层:①肺表面活性物质;②液体分子;③肺泡上皮细胞;④间质(弹力纤维和胶原纤维);⑤毛细血管基膜;⑥毛细血管内皮细胞。6层结构的总厚度仅为0.2~1μm,通透性大,气体容易扩散通过。
2.肺泡表面活性物质(alveolar surfactant) 是由肺泡Ⅱ型细胞分泌的一种复杂的脂蛋白,主要成分是二棕榈酰卵磷脂。肺泡表面活性物质的主要功能是:
(1)降低肺泡的表面张力
(2)维持肺泡内压的相对稳定
(3)阻止肺泡积液
二、肺通气原理
气体进出肺取决于两方面因素的相互作用:一是推动气体流动的动力;二是阻止其流动的阻力。
(-)肺通气的动力
气体之所以能够进出肺是由于在肺内与大气之间存在着压力差。肺回缩时,肺容积减小,肺内压大于大气压,肺内气体排出体外;肺扩张时,肺容积增大,肺内压低于大气压,空气进入肺内。但是肺本身是一个富有弹性的器官,不能自行张缩,而是靠呼吸肌的活动被动牵拉实现张缩的。
1.呼吸运动
(1)吸气运动吸气总是主动过程。平和呼吸时,肋间外肌收缩,肋骨向前向外移动,胸廓的上下左右径增大;隔肌收缩时,膈向后移动,胸廓前后径增大。胸腔容积增大肺被动牵引而扩张,肺容积增大,肺内压下降低于大气压,空气趁虚而进入肺内,引起吸气(inspiration)。
(2)呼气运动平和呼吸时,吸气终末,肋间外肌和膈肌由收缩转为舒张,肋和隔回位,胸腔容积减小,肺失去牵引力由自身的弹性和表面张力而回缩,肺容积减小,肺内压升高,高于大气压时,肺内气体压出体外,引起呼气(expiration)。呼气运动是被动的。
2.呼吸频率及呼吸式一分钟内呼或吸的次数为呼吸频率(respiratory frequency)。
哺乳动物呼吸式(breathing pattern)有三种类型:
胸式呼吸(thoracic breathing):吸气时以肋间外肌收缩为主,胸壁起伏明显
腹式呼吸(abdominal breathing):吸气时以隔肌收缩为主,腹部起伏明显
胸腹式呼吸(混合式呼吸)(combined breathing):吸气时肋间外肌与隔肌都参与,胸壁和腹壁的运动都比较明显,
3.肺内压与胸内压肺内压(intrapulmonary pressure)是指肺泡内的压力。吸气初,肺容积增大,肺内压下降,低于大气压,空气进入肺泡,肺内压也逐渐升高;吸气未,肺内压与大气压相等,气流也就停止。呼气初,肺容积减小,肺内压暂时升高,超过大气压,肺内气体便流出肺,肺内气体逐渐减少,肺内压下降;至呼气未,肺内压降至与大气压相等。
由上述可见,在呼吸过程中正是由于肺内压的周期性交替升降,造成肺内压与大气压之间的压差,这一压差成为推动气体进出肺的直接动力。
胸内负压是如何形成的?胸内压实际上是通过胸膜脏层作用于胸膜腔间接形成的。胸膜壁层的外表面有坚厚的胸廊组织支持,胸壁上的大气压力不会影响胸膜腔。而胸膜脏层却受到两方面力的影响,一是肺内压,即大气所加的压力,使肺泡扩张;一是肺的回缩力,使肺泡缩小。因此,胸膜腔内的压力是上述两种方向相反的力的代数和,即:
胸内压=肺内压-肺回缩力
在吸气之末和呼气之末,肺内压等于大气压,因此:
胸内压=大气压-肺回缩力
如以一个大气压作为生理零位标准,则
胸内压=-肺回缩力
由此可见,胸内负压是由肺的回缩力造成的。吸气时肺扩张,回缩力增大,负压也增大;呼气时相反,负压减小。如马在平和呼吸时,吸气末胸内负压值为 2.12 kPa(-16 mmHg)呼气末为 0.79 kPa(-6mmHg)。
胸内压负压具有重要的生理意义。首先,胸内负压是肺扩张的重要条件,由于胸膜腔与大气隔绝,处于密闭状态,因而对肺有牵拉作用,使肺泡保持充盈气体的膨隆状态,能持续地与周围血液进行气体交换,不致于在呼气之末肺泡塌闭无气体而中断气体交换;其次,胸内负压对胸腔内的其它器官有明显的影响。如吸气时,胸内压降得更低,引起腔静脉和胸导管扩张,促进静脉血和淋巴回流。胸内负压还可使胸部食管扩张,食管内压下降,因此在呕吐和反刍逆呕时,均表现出强烈的吸气动作。
(二)肺通气的阻力
肺通气的阻力来自于两方面:①肺与胸廓的回位力,称弹性阻力;②呼吸道气流阻力(气体与呼吸道管壁之间,气体分子之间所产生的摩擦阻力)以及肺和胸廓活动时,有关组织之间的粘滞阻力,总称为非弹性阻力。
1.弹性阻力和顺应性弹性组织在发生变形时,要产生阻止变形恢复原位的力,称弹性阻力。肺与胸廓都属于弹性组织,无论是扩张还是收缩,只要发生变形,都会产生弹性阻力使其回位。因此,呼吸肌必需收缩作功,只有克服弹性阻力,胸廓和肺才能张缩,才能实现通气机能。
顺应性(lung compliance)是指在外力作用下,弹性组织的可扩展性,容易扩展者,顺应性大;反之,则顺应性小。可见顺应性(C)与弹性阻力(R)成反变关系:
因此,顺应性是衡量弹性阻力大小的指标。而顺应性(C)则可用单位压力变化所引起的容积变化来衡量。
肺的弹性阻力是由肺的弹性纤维变形时产生的回位力与肺泡的表面张力共同组成的。若肺组织纤维化或由于肺炎、肺血栓等疾病而引起肺表面活性物质分泌减少,表面张力增大,都可导致肺的弹性阻力增大,肺不易扩张,也即肺的顺应性变小。即:
跨肺压是肺内压与胸内压之差。猫肺静态顺应性曲线表明,跨肺压增大,肺容积也相应增大,曲线中间段压力与容积两者几乎呈直线关系,说明在正常生理范围内顺应性数值是相对稳定的。
胸廓的弹性阻力来自胸部肌肉、腱、结缔组织、腹壁肌以及腹腔内脏等弹性组织。胸廓畸形、肥胖等因素均可增大弹性阻力,胸廓顺应性减小。
呼吸系统内肺的弹性阻力与胸廓的弹性阻力相互串联,总的弹性阻力等于肺与胸廓弹性阻力之和。由于弹性阻力与顺应性呈倒数关系,故总弹性阻力又为肺与胸廓顺应性倒数之和。在呼吸过程中胸廓的弹性阻力发生向量变化,因此总的弹性阻力实际是肺与胸廓弹性阻力的代数和。
正常肺始终处于扩张状态,总有弹性回缩力。胸廓的弹性回缩力的方向又随其扩张与回缩程度发生向量变化。肺容量小于机能余气量时(相当于深呼气),胸廓的弹性回缩力向外而且大于肺向内的弹性阻力;肺容量为机能余气量时(平和呼气),肺的弹性阻力与胸廓的弹性阻力大小相等,但方向相反,相互抵消;肺容量大于机能余气量时(平和吸气)胸廓在自然位置,不表现弹性阻力,而只有肺的回缩力;肺容量接近肺总容量时(深吸气),肺与胸廊的弹性阻力都向内,共同组成肺扩张阻力。
2.非弹性阻力非弹性阻力包括惯性阻力、粘滞阻力和气道阻力。惯性阻力是气流在发动、变速、换向时因气流和组织惯性所产生的阻止气体流动的因素。平和呼吸时,惯性阻力小,可忽略不计。粘滞阻力来自呼吸时组织相对位移所发生的摩擦。气道阻力来自气体流经呼吸道时,气体分子之间以及气体分子与气道壁之间的摩擦,这是非弹性阻力的主要成分,约占80%~90%。非弹性阻力是在气体流动时产生的,并随气流速度加快而增加。下面主要讨论气道阻力。
气流阻力受气流流速、气流形成和呼吸道管径大小的影响。流速快,阻力大;流速慢,阻力小。气流形式有层流和湍流,层流阻力小,湍流阻力大。气流太快和管道不规则,容易发生湍流。气道管径大小是影响气道阻力的重要因素。管径小,阻力大(R∝1/r4)。吸气时,胸内压下降,气道扩张,阻力减小;呼气时则反之。气管平滑肌受交感与迷走神经支配,交感神经兴奋,气管扩张,阻力减小;迷走神经兴奋,气管平滑肌收缩,管径缩小,阻力增大。
呼吸肌收缩作功,安静时2/3用来克服弹性阻力,l/3用来克服气流阻力。
- 肺容量与肺通气量
(一)肺容量
肺容量是指肺内容纳气体的量,随进出肺的气体量而变化。肺容量由以下几部分组成。
1.肺活量 用力吸气后再用力呼气,所能呼出的气体量。即潮气量、补吸气量与补呼气量之和为肺活量(vital capacity)。肺活量反映了一次通气时的最大能力,在一定程度上可作为肺通气机能的指标。
(1)潮气量平静呼吸(eupnea)时,每次吸入或呼出的气体量,为潮气量(tidal volume,TV)。运动或使役时增大。
(2)补吸气量平和吸气未,再尽力吸气,多吸入的气体量为补吸气量(inspiratory reserve volume)。
(3)补呼气量平和呼气末,再尽力呼气,多呼出的气体量为补呼气量(expiratory reserve volume)。
2.余气量补呼气后肺内残留的气体量为余气量(residual volume)。
3.机能余气量平和呼气后肺内残留的气体量为机能余气量(functional residual capacity)。即补呼气量与余气量之和。机能余气量约相当于潮气量的4~5倍。机能余气量的生理意义在于:每次从外界吸入或自肺循环进入肺泡的气体,首先被机能余气量稀释,缓冲了肺泡中氧和二氧化碳分压(PO2和PCO2)的急剧变化。由于机能余气量的稀释作用,吸气时肺内 PO2不致突然升得太高,PCO2也不致降得太低;呼气时肺内PO2不致于降得太低,PCO2也不致升得太高。这样,肺泡气和动脉血液中的PO2和PCO2就不会随着呼吸发生大幅度波动。
4.肺总容量肺所容纳的最大气体量为肺总容量(total lung capacity)。即肺活量与余气量之和。各种动物的肺容量不同,如马为4OL。马的潮气量为6 L、补吸气量与补呼气量各为12L,余气量为10L。
(二)肺通气量
1.每分通气量每分通气量(minute ventilation volume)是指每分钟进或出肺的气体总量。
每分通气量 =潮气量×呼吸频率
健康动物的潮气量和呼吸频率随着机体代谢水平而变化。代谢水平增高,如运动或使役时,呼吸频率和潮气量都会增大,每分通气量也增大。如马在休息时,每分通气量为35~45 L,平地步行时为 80~150 L,负重时为150~250 L,挽拽时为 300~450 L。
每分钟肺能够吸入或呼出的最大气体量,称为肺的最大通气量。健康动物的最大通气量可比平和呼吸时的每分通气量大10倍多。肺的最大通气量反映了肺在每分钟的最大通气能力,它是比肺活量更能客观地反映肺通气机能的指标之一。
每分最大通气量与每分通气量之差可表明肺通气量的储备力量。
通气贮备量是反映呼吸机能的良好指标,并可以此判断通气储备能力。
2.肺泡通气量(alveolar ventilation)每次吸入的气体,一部分停留在呼吸性细支气管以上部位的呼吸道内,这部分气体不能参与肺泡间的气体交换,称为解剖无效腔或死腔。进入肺泡内的气体,也可能由于血液在肺内分布不均而未能与血液进行气体交换。未能发生气体交换的这部分肺泡容量称肺泡无效腔。肺泡无效腔与解剖无效腔一起合称为生理无效腔。健康动物的肺泡无效腔接近于0,因此,生理无效腔几乎与解剖无效腔相等。由于无效腔的存在,每次吸入的新鲜空气,一部分停留在无效腔内,另一部分进入肺泡。可见肺泡通气量才是真正的有效通气量。肺泡通气量按下式计算:
肺泡通气量=(潮气量-无效腔量)×呼吸频率
每分通气量与每分肺泡通气量之差,除以呼吸频率所得的商,即生理无效腔量。生理无效腔与解剖无效腔气量之差,可反映非功能性肺泡容量。
第二节 气体交换及运输
在呼吸过程中气体交换发生在两个部位:一是肺与血液间的气体交换,称肺换气;另一是组织与血液间的气体交换,称组织换气。经肺换气与组织换气进入血液中的O2与CO2,经血液循环分别运送到组织和肺,这就是气体交换与运输。
- 气体交换
(一)气体交换原理
气体分子不停地进行着无定向的运动,其结果是气体分子从高分压向低分压扩散。这就是气体交换的原理。
(二)肺和组织内的气体交换
1.肺换气气体总是由分压高的一侧透过呼吸膜向分压低的另一侧扩散。因此,肺泡气中的O2透过呼吸膜扩散进入毛细血管内,而血中的CO2透过呼吸膜扩散进入肺泡内。
2.组织换气依据气体由高分压向低分压扩散的规律,组织中的CO2进入血液,而血液中的O2进入组织。毛细血管中的动脉血,边流动边进行气体交换,逐渐变成静脉血。
(三)影响肺内气体交换的因素
1.分压差、溶解度和分子量气体扩散率与气体分压差、溶解度成正比,与分子量平方根成反比。
CO2的溶解度比O2的溶解度大得多(24:1);在同样分压下,CO2的扩散速度要比O2快得多,约为O2的20倍。通常情况下,肺换气不足时往往缺O2显著,而CO2储留不明显。
2.呼吸膜面积与厚度单位时间内气体的扩散量与呼吸膜面积成正比,与厚度成反比。
3.通气/血流比值 (ventilation/perfusion ratio)是指每分肺泡通气量(VA)和每分肺血流量(Q)之间的比值(VA/Q)。如果比值增大,则表明部分肺泡不能与血液中气体充分交换,即增大了肺泡无效腔;比值减小,则表明通气不良,血流过剩,部分血液流经通气不良的肺泡,混合静脉血中的气体未得到充分更新,未能成为动脉血就流回了心脏,犹如发生了动—静脉短路一样。由此可见无论VA/Q增大,还是减小,两者都妨碍了有效气体交换,导致血液缺O2或CO2储留,尤其是缺O2。
- 气体运输
从肺泡扩散入血液的O2必须通过血液循环运送到各组织,从组织扩散入血液的CO2也必须由血液循环运输到肺泡。
O2与CO2都以物理溶解和化学结合两种形式存在于血液中,但以溶解形式存在的极少,绝大部分呈化学结合形式。
体内血液中的 O2和 C02的物理溶解和化学结合状态时刻保持着动态平衡。在肺或组织进行气体交换时,进入血液中的 O2和 CO2都是先溶解,提高分压后再结合。O2和 CO2从血液释放时,也是溶解的先逸出,分压下降,结合的再分离出来补充所失去的溶解的气体。
(-)氧的运输
血液中的O2主要是与红细胞内的血红蛋白(Hb)结合,以氧合血红蛋白(HbO2)的形式运输,占98. 4%;溶解O2甚微,仅占1.6%。
1.血红蛋白与O2的可逆结合血红蛋白与O2结合有下列特征:
①反应快、可逆、不需酶催化,P O2高时(肺部),血红蛋白与O2结合形成氧合血红蛋白(HbO2);P O2降低时(组织),氧合血红蛋白迅速解离,释放O2。
②血红蛋白与O2结合,其中铁仍为二价,所以该反应不是氧化而是氧合(oxygenation)。
③只有在血红素的Fe2+和珠蛋白的链结合的情况下,才具有运输O2机能,单独的血红素不是有效的氧载体。
④l分子血红蛋白可与4分子O2结合。
100 ml血液中Hb所能结合的最大氧量称Hb氧容量(血氧容量)。Hb实际结合的O2量称 Hb的氧含量(血氧含量)。Hb氧含量与氧容量的百分比为Hb氧饱和度。
2.氧分压与氧离曲线氧离曲线或氧合血红蛋白解离曲线(oxygen dissociation curve)是表示PO2与Hb氧饱和度的关系曲线。该曲线即表示不同PO2下,O2与Hb解离情况,同样也反映了不同PO2时O2与 Hb的结合情况。
氧离曲线上段相当于PO2值8~13.33 kPa(60~100 mmHg)范围,这段曲线较平坦,表明PO2的变化对Hb氧饱和度影响不大,Hb氧饱和度在90%以上。在这个范畴内,即使PO2有所下降,只要PO2不低于 60 mmHg, Hb氧饱和度仍能保持在90%以上,血液仍可携带足够的氧,不致发生明显的低O2症。
氧离曲线下段相当于PO25 .33~1.33 kPa(40~10 mmHg)。这段曲线陡直,是HbO2释放O2的部分,也即 PO2稍有下降,Hb氧饱和度就会有较大幅度下降(相当于组织部位的P O2波动范围),有较多的O2释放出来供组织活动需要。
氧离曲线呈“S”形。
Hb与O2的结合和解离可受多种因素的影响:
(1) pH值和CO2浓度的影响血液中的pH值越低或 PCO2越高,Hb氧饱和度下降越明显,氧离曲线右移;反之则氧饱和度升高。
(2)温度的影响温度增高可使氧离曲线右移。
(3)2,3—二磷酸甘油酸(2,3—DPG)当血液的PO2降低时,红细胞内无氧酵解增强,致使2,3—DPG产生增多。 2,3—DPG在调节Hb和O2的亲和力中起重要作用。2,3—DPG浓度升高,Hb和O2亲和力下降,氧离曲线右移;反之2,3—DPG浓度降低, Hb和O2的亲和力则增加,曲线左移。
(4)Hb自身性质的影响 当Hb中Fe2+氧化成Fe3+时,则会失去运输氧气的能力。
(二)二氧化碳的运输
CO2有两种存在形式:溶解形式和结合形式
一部分CO2进入红细胞内,与Hb的-NH2结合,形成氨基甲酸血红蛋白(HbNHCOOH),亦称碳酸血红蛋白(HbCO2)。这一反应很迅速,无需酶参与。
组织中的CO2扩散进入血液后透过红细胞膜进入红细胞内,由于红细胞内含有较高浓度的碳酸酐酶,在其作用下,H2O和 CO2迅速生成H2CO3,并迅速分解成为 H+和HCO3-。在生成H2CO3的同时,红细胞内的氧合血红蛋白钾盐(KHbO2),由于组织内的PO2低而放出O2,生成脱氧血红蛋白钾盐(KHb)。KHb酸性较弱,它所结合的钾容易被H2CO3中的H+所置换,生成HHb和KHCO3,即:
CO2不断进入红细胞,使HCO3-含量逐渐增多,当超过血浆中HCO3-的含量时,HCO3-透过红细胞膜扩散进入血浆,并与血浆中的 Na+结合生成 NaHCO3。在HCO3-扩散入血浆的过程中,又有等量的Cl-从血浆扩散入红细胞,以维持红细胞内外正负离子的静电平衡。这种Cl-与HCO3-的交换现象,称为氯转移(chloride shift)。这样HCO3-不致在红细胞内蓄积,以利组织中的CO2不断进入血液。生成的 KHCO3(红细胞)和 NaHCO3(血浆中)经血液循环运至肺部。
当静脉血流经肺泡时,由于肺泡中的PCO2比静脉血低,同时红细胞中的还原血红蛋白(HHb)大部分与氧结合生成氧合血红蛋白(HbO2),氧合血红蛋白又与 KHCO3作用生成H2CO3。红细胞内的H2CO3在碳酸酐酶催化下,分解为CO2和H2O,CO2扩散进入血浆,进而扩散到肺泡气中,经肺呼出体外。
这样,红细胞内的H2CO3逐步降低,于是血浆中的NaHCO3分解,HCO3-进入红细胞内,与此同时红细胞内的Cl-又返回血浆,进行反向的氯转移。
第三节 呼吸运动的调节
- 神经调节
(一)呼吸中枢
呼吸中枢是指中枢神经系统内产生和调节呼吸运动的神经细胞群。
1.脊髓 脊髓是呼吸运动的初级中枢,联系上位呼吸中枢并通过位于腹角的运动神经元,支配呼吸肌的运动。在呼吸反射活动中,脊髓也起某些整合作用。
2.延髓 基本的呼吸节律产生于延髓,延髓的呼吸神经元主要集中分布在延髓的背侧和腹侧的两组神经核团内,分别称为背侧呼吸组(dorsal respiratory group,DRG)和腹侧呼吸组(ventral respiratory group,VRG)。背侧呼吸组呼吸神经元主要集中于孤束核的外侧,包括吸气神经元(IN)和呼气神经元(EN)。)
3.脑桥脑桥上部呼吸神经元相对集中于臂旁内侧核与相邻的K?lliker-Fuse(KF)核,合称PBKF核群。该核群和延髓的呼吸神经核团之间有双向联系,其作用是限制吸气,促使吸气向呼气转换。
(二)呼吸的反射性调节
呼吸活动可受机体内外环境各种刺激的影响,如伤害性刺激、冷刺激、血压的骤然变化等都可使呼吸发生变化。重要的反射如下:
1.肺牵张反射(pulmonary stretch reflex)由肺扩张或肺缩小引起的吸气抑制或兴奋的反射称黑—伯二氏反射(Hering-Breuer reflex),又称肺牵张反射。,呼吸加深变慢。
2.呼吸肌的本体感受性反射此反射是指呼吸肌的本体感受器肌梭(属机械感受器)受牵张刺激时,上传冲动而引起呼吸肌反射性收缩加强。
3.防御性呼吸反射当鼻腔、咽、喉、气管与支气管的粘膜受到机械或化学刺激时,则会引起防御性反射。此反射具有清除刺激物,以防异物进入肺泡的作用。
- 化学因素对呼吸的调节
调节呼吸的化学因素是指动脉血液或脑脊液中的O2、CO2和H+。当血液或脑脊液中的CO2、H+浓度升高,O2浓度降低时,刺激化学感受器通过调节呼吸,排出体内过多的CO2、H+,摄入O2,以维持血液与脑脊液中CO2、O2、H+浓度的相对恒定。
(一)化学感受器(chemoreceptor)
1.中枢化学感受器位于延髓腹外侧浅表部位,左右对称。
2.外周化学感受器位于颈动脉窦与主动脉弓附近,分别称为颈动脉体和主动脉体。外周化学感受器对血液中缺O2和H+增高很敏感。
(二)二氧化碳对呼吸的影响
血液中CO2含量轻度变化则对呼吸有显著的影响。当动脉血中PCO2增高可使肺通气量增大,加快CO2的排出,维持血中的CO2含量的相对恒定。
(三)低氧对呼吸的影响
吸入空气中PO2在一定范围内下降可以引起呼吸增强。
(四)氢离子对呼吸的影响
动脉血中H+增加,呼吸加深加快;H+降低,呼吸受到抑制。
第六章 消化生理
消化(digestion):食物在消化道内被分解为可吸收的小分子物质的过程。
吸收(absorption):食物经过消化后的产物、水、盐类等,透过消化道粘膜,进入血液和淋巴循环的过程。
消化和吸收是两个相辅相成、紧密联系的过程。不能被消化吸收的食物残渣,最终以粪便的形式排出体外。
第一节概述
一、消化的方式
1.机械性消化是指通过消化道肌肉的舒缩活动,将食物磨碎,并使之与消化液充分混合,以及将食糜不断地向消化道远端推送,最终将消化吸收后的饲料残渣排出体外。
2.化学性消化是指消化腺所分泌的各种消化酶和植物饲料本身的酶将饲料中的蛋白质、脂肪和糖类分解成为小分子物质的过程。
3.生物学消化是指由栖居在畜禽消化道内的微生物对饲料进行发酵的过程。此种消化方式对饲料中纤维素、半纤维素、果胶等高分子糖类的消化具有极为重要的意义。
上述三种消化过程既具有明显的阶段性,又相互联系、相互影响和同时进行。
二、消化道平滑肌的特性
(一)消化道平滑肌的基本特点
1.兴奋性较低、收缩缓慢消化道平滑肌的兴奋性比骨骼肌低。收缩的潜伏期、收缩期和舒张期比骨骼肌所占的时间要长的多,而且变异很大,即收缩缓慢。
2.自动节律性离体的胃肠道平滑肌,在适宜的环境中,仍能进行良好的节律性运动,但其收缩很缓慢,节律性也远不如心肌规则。它起源于平滑肌本身,在体条件下也受神经系统的调节。
3.较大的展长性可根据需要伸展,最长时可为原来的2-3倍。
4.紧张性经常保持在一种微弱的持续收缩状态,即便离体后,只要放在适意的溶液中,仍能自动进行持续的收缩,这种特性叫紧张性。
5.对化学、温度和牵张的刺激较敏感如肾上腺素可增高静息电位,使肌纤维的兴奋性降低,张力减弱,Ach却降低静息电位,加强肌纤维的兴奋和张力。食麋进入某段引起牵拉,使平滑肌膜去极化,增强肌肉的收缩。温度剧变或缺O2时,也可降低静息电位,引起肌肉收缩增强。
(二)消化道平滑肌的电位变化
1.静息电位较小,约-60~-50mV,电位的产生主要由K+的平衡电位形成。
2.基本电节律(basal electric rhythm,BER)消化道平滑肌细胞可自发产生节律性的去极化。电生理研究表明:在安静状态下,用微电极可在胃肠道纵行肌细胞静息电位基础上记录到一种缓慢的、大小不等的、节律性去极化波,由于其发生频率较慢而被称为慢波电位,又称基本电节律,其波幅变动在20~30mV之间,持续1~4秒,频率随消化道不同部位而异。目前认为:慢波的产生可能与细胞膜上的生电性钠泵活动的周期性活动有关。另有人认为是Na+内流,K+外流形成的。
3.峰电位是迅速而短暂的去极化波,但一般都是发生在基础电节律之上,此现象称为峰簇。
慢波引起快波,快波引起收缩。
平滑肌产生的慢波电位本身并不一定引起动作电位和平滑肌收缩。如果在神经或体液因素的影响下,慢波电位超过一定临界值时,就可能在慢波的脊上触发一个或多个动作电位,随之出现平滑肌收缩。每个慢波上所出现动作电位的数目越多,肌肉收缩的幅度就越大。
三 胃肠道功能的调节
(一)胃肠道的神经支配及作用
1.交感神经和副交感神经胃肠道受交感神经和副交感神经的双重支配。其中副交感神经的作用是主要的。
2.内在神经丛也叫壁内神经丛(intrinsic plexus),分布在从食管中段到肛门的绝大部分消化管壁内。
(二)胃肠激素及作用
1.胃肠激素胃肠道的内分泌细胞,分泌的多种激素和激素类物质,统称为胃肠激素(gastrointestinal hormone)。这些激素在化学结构上都是由氨基酸残基组成的肽类,分子量大约在5000以内。它们与神经系统共同调节着消化器官的运动、分泌、吸收等活动。
胃肠道粘膜的内分泌细胞的数量超过了体内所有内分泌腺中内分泌细胞的总和,是体内最大的、最复杂的内分泌器官这些内分泌细胞都具有摄取胺前体,进行脱羧而产生肽类激素或活性胺的能力,具有这种能力的细胞统称为APUD(amine precursor uptake and decarboxylation)细胞。
除胃肠和胰腺的内分泌细胞外,神经系统、甲状腺、肾上腺髓质、垂体等组织中也含有APUD细胞。它们分泌的激素主要通过血液循环运送到靶细胞起作用,但也有一些胃肠激素释放后并不进入血液循环,而是通过细胞外液弥散至邻近的靶细胞,此种传递局部信息的方式称为旁分泌。
2.胃肠激素的作用胃肠激素的主要作用是:
(1)调节消化腺的分泌和消化道的运动
(2)调节其它激素的释放现已证明,食物消化时,从胃肠释放的抑胃肽(gastric inhibitorypolypeptide,GIP)有很强的刺激胰岛素分泌的作用。
(3)营养作用一些胃肠激素具有刺激胃肠道组织的代谢和促生长的作用,称为营养作用(trophic action)。例如,胃泌素能刺激胃的泌酸部位粘膜和十二指肠粘膜的蛋白质、RNA和DNA的合成,从而促进其生长。给动物长期注射五肽胃泌素(一种人工合成的胃泌素),可引起壁细胞增生。胆囊收缩素能引起胰腺内DNA、RNA和蛋白质的合成增加,促进胰腺外分泌组织的生长。
(三)脑—肠肽的概念
近年来的研究证实,一些产生于胃肠道的肽,不仅存在于胃肠道,也存在于中枢神经系统内;而原来认为只存在于中枢神经系统的神经肽,也在消化道中发现。这种双重分布的肽统称为脑—肠肽(brain-gut peptide)。
已知的脑—肠肽约有20余种。这些肽的双重分布具有很重要的生理意义。如胆囊收缩素在外周对胰酶分泌和胆汁排放具有调节作用,在中枢对摄食具有抑制作用,提示脑内及胃肠内的胆囊收缩素在消化和吸收中具有协调作用。
第二节口腔内的消化
口腔消化从食物进入口腔后开始,经过咀嚼,混入唾液,形成食团后吞咽入胃。
一、采食和饮水
(一)采食和饮水的方式
各种动物各有特点,由采食习性决定。
狗、猫和其它肉食动物通常用前肢按住食物,用门齿和犬齿咬断食物,并依靠头部和颈的运动将食物送入口内。
牛的主要采食器官是舌。
马、驴主要靠唇和门齿采食。
绵羊和山羊的采食方法与马大致相同。绵羊上唇有裂隙,便于啃食很短的牧草。
猪用鼻突掘地寻找食物,并靠尖形的下唇和舌将食物送入口内。
饮水时,猫和狗将舌浸入水中,卷成匙状,将水送入口中。其它家畜一般先把上下唇合拢,中间留一小缝,伸入水中,然后下颌下降,同时舌向咽部后移,使口腔内形成负压,把水吸入口腔。仔畜吮乳也是靠口腔壁肌肉和舌肌收缩,使口腔形成负压来完成。
(二)采食的调节
1.采食中枢随意采食主要受神经系统的调控,体液因素(包括激素和代谢产物)也参与作用。
在实验室动物研究表明,在下丘脑存在食物中枢,有摄食中枢和饱中枢组成。摄食中枢又称饥饿中枢,位于下丘脑左右两侧的外侧区,刺激这个中枢,可使刚吃饱的动物恢复摄食活动,破坏后可导致动物厌食,甚至饿死。饱觉中枢位于下丘脑两侧的腹内侧核,刺激这个中枢可使动物停止摄食,破坏则出现暴食,形成肥胖。这种情况也在猪、鸡及山羊实验获得证实。
2.反射调节
采食还受反射性调节(口、咽刺激、胃肠的机械刺激,胃肠道渗透压核化学效应、吸收后的渗透压和化学效应)、激素及神经递质(NE、Ach、GABA、cck-PE、内啡肽)等调节。
二、唾液分泌
(一)唾液腺
浆液型:腮腺、下颌腺
粘液型:上颌腺、口腔粘膜的小腺体
混合型:舌下腺
唾液是腮腺(parotid gland)、颌下腺(submaxillary gland)和舌下腺(sublingual gland)三对主要唾液腺颌口腔粘膜中许多小腺体分泌地混合液。
(二)唾液腺分泌的研究方法
组织培养(细胞培养法)
慢性瘘管法唾液碗:瘤胃贲门处放置
体外吻合瘘
(三)唾液的性质和成分及作用
唾液为无色透明的粘稠液体,呈弱碱性,
水分(99.4%)
唾液无机物(0.6%K Na Ca Mg 氯化物 磷酸盐 碳酸氢盐等)
有机物:粘蛋白颌其它蛋白质,酶α唾液淀粉酶
唾液脂肪酶
溶菌酶
血型物质
作用:
(1)湿润口腔和饲料,便于咀嚼和吞咽。
(2)溶解食物中的各种可溶性物质,
(3)唾液淀粉酶在接近中性的条件下,可水解淀粉为麦芽糖。
(4)唾液中的粘蛋白有润滑作用,便于形成食团,利于食团的吞咽和在食道中的运输。
(5)反刍动物唾液中高浓度的碳酸氢盐和磷酸盐具有强大的缓冲能力,能中和瘤胃内微生物发酵所产生的有机酸,借以维持瘤胃内适宜的酸碱度,保证微生物正常活动。
(6)唾液能冲淡和洗去口腔中的饲料残渣和异物,洁净口腔;狗等肉食动物唾液中的溶菌酶具有杀菌作用。
(7)保持口腔的碱性环境,使饲料中的碱性酶免受破坏,在其进入胃的初期仍发挥消化作用。
(8)某些动物(如牛、狗)的汗腺不发达,在高温季节可分泌大量稀薄唾液,其中水分的蒸发有助于散热。
(9)反刍动物可随唾液分泌大量的尿素进入瘤胃,参与机体的尿素再循环,减少氮的损失。
(四)唾液分泌的调节
1、神经调节
唾液分泌完全受神经的反射性调节,包括条件反射和非条件反射。
食物的形、色、味以及采食时的周围环境等各种信号,可以建立条件反射而引起的唾液分泌。非条件反射性唾液分泌是指食物对口腔的机械、化学、温度等刺激引起口腔粘膜及舌部的感受器兴奋,
支配唾液腺的神经为交感神经和副交感神经,副交感神经兴奋,引起颌下腺和腮腺大量分泌;刺激交感神经地反应,各种动物不一样,如狗:颌下腺分泌,不引起腮腺分泌;猫:颌下腺分泌的同时,腮腺少量分泌。
2、体液调节
主要为激素调节,手术时防唾液分泌过多注射阿托品;醛固酮调节Na+/K+
3、其它因素
唾液分泌量取决于食物地含水量、味道、成分、动物种类、年龄差别等。
反刍动物瘤胃内地压力颌化学感受器受到刺激,引起腮腺不间断地分泌。
急性病、疼痛、脱水和强烈地精神刺激等,见唾液分泌减少。
三、咀嚼
饲料入口后,被送到上下颌臼齿间,在咀嚼肌的收缩和舌、颊部的配合运动,食物被粉碎,并混合唾液。
咀嚼的意义:(1)机械地将饲料粉碎,并破坏其细胞地纤维膜,使饲料地消化面积增加,有利于消化。(2)使粉碎后地饲料与唾液混合,并形成食团便于吞咽。(3)反射性地引起消化腺地活动颌胃肠运动,给以后地消化过程准备有利条件。
四、吞咽
吞咽(deglutition)是由口腔、舌、咽和食管肌肉共同参与的一系列复杂的反射性协调活动,是食团从口腔进入胃的过程。
吞咽动作
食物经咀嚼形成食团后,随即由于颊部颌舌地运动将食团送达咽部,刺激该部的感受器,反射性地引起软腭上举并关闭鼻咽孔,阻断口腔与鼻腔的通路。同时会厌软骨翻转,盖住喉口,呼吸暂停;此时,口腔和咽部形成密闭室,同时食管口舒张,咽肌收缩,将食团迅速挤入食管,又供食管蠕动送至贲门进入胃内,这个过程形成吞咽动作。
第三节 单胃内的消化
一、胃的粘膜结构
胃粘膜一般分为:贲门腺区、胃底腺区和幽门腺区和与食管相接地无腺部(食管膨大部)。
马无腺部较大,猪地则很小。
贲门腺分泌盐酸
胃底腺主细胞:胃蛋白酶、凝乳酶、脂肪酶
壁细胞:盐酸
粘液细胞:粘蛋白
幽门腺分泌粘液
无腺部覆以扁平细胞
整个胃粘膜分布粘液细胞,分泌粘液形成保护层。
二 胃液的分泌
1.胃消化酶(pepsin)
(1)胃蛋白酶:一种蛋白水解酶,由主C分泌。蛋白质→月示 +月东
(2)胃脂肪酶:分解脂肪,主C分泌。幼畜较多,活性弱。乳化脂肪→甘油+脂肪酸
(3)凝乳酶:使乳凝固,犊牛含量高酪蛋白原→酪蛋白酪→蛋白钙
2.盐酸通常所说的胃酸,即为盐酸,由壁细胞分泌。小部分与粘液中的有机物结合,称为结合酸;大部分以游离方式存在,称为游离酸;二者合称为总酸。
盐酸主要具有以下生理作用:(1)激活胃蛋白酶原,使它转变成有活性的胃蛋白酶,并为其提供适宜的酸性环境。(2)使蛋白质膨胀变性,便于被胃蛋白酶水解。(3)抑制和杀灭随饲料进入胃内的微生物,维持胃和小肠的无菌状态。(4)盐酸进入小肠后能刺激促胰液素的释放,从而促进胰液、胆汁和小肠液的分泌。(5)盐酸所造成的酸性环境有助于铁和钙的吸收。
3.粘液
主要成分为大分子的糖蛋白。
粘液不溶性粘液:表面上皮分泌,机械刺激时分泌。
可溶性粘液:粘液细胞分泌,迷走神经兴奋时分泌。
作用:(1)润滑和保护作用,使胃粘膜不受饲料中坚硬物质的机械损伤。
(2)防止自消化作用,粘液呈弱碱性反应,可降低胃酸的酸度,减弱胃蛋白酶的活性,从而防止酸颌胃蛋白酶对粘膜的消化作用。
三 胃液分泌的调节
受神经和体液双重调节,胃消化过程中的胃液分为头期、胃期和肠期。
- 头期由进食动作引起的胃液分泌,可用“假饲”实验证实、食管瘘管。
假饲→潜伏期→胃液分泌 系非条件反射
传入神经→延髓→丘脑下部及大脑皮质等的分泌中枢→迷走神经兴奋(Ach释放)→胃液分泌
特点是:持续时间长、分泌量大、酸度高、胃蛋白酶含量高、消化力强。
- 胃期
头期只是胃液分泌的开始,食物入胃后,还可继续刺激引起胃液分泌。
- 反射性分泌食物的硬度和容积刺激胃壁的压力感受器,反射性引起胃液分泌,传出和传入神经都在迷走神经内。
- 体液性分泌反射性分泌之后,胃液还可继续分泌,这是由于胃粘膜G细胞分泌促胃液素(胃泌素 Gastrin),经血循道胃腺,引起胃液分泌加强。
Gastrin为17个AA的直链多肽,犬胃泌素(G-34),较小胃泌素(G-17),最小胃泌素(G-14).
引起Gastrin的分泌因素:(1)迷走神经兴奋,(2)机械刺激(扩张)化学刺激(酒精、蛋白分解产物),(3)碱化幽门窦粘膜促进其分泌。
G作用:刺激壁细胞分泌盐酸,刺激小肠粘膜,也能产生促胃液素,盐酸进入十二指肠会抑制胃液分泌。
3、肠期胃内食麋进入十二指肠,刺激小肠粘膜,也能产生促胃液素,盐酸进入十二指肠会抑制胃液分泌。
肠期胃液分泌的特点是:分泌量少,大约只有采食后胃液分泌总量的1/10,消化力低。
四、胃的运动及排空
胃空虚时,胃壁紧缩,采食时由于食团刺激咽部颌食管,反射性的通过迷走神经引起胃底和胃体部的肌肉舒张,这一反应称为容受性舒张。
空腹时胃内压力等于腹内压,由于有容受性舒张,进食后胃容量增加时,胃内压力增加很小,从而使胃完成容受和储存食物的机能。
1、胃在消化期的运动
食物进入胃后,呈明显的分层排列,且这种分层在胃内保持较长时间,使胃液不能迅速浸透胃内所有食团。此期胃运动有两种形式:
一是紧张性收缩:是以平滑肌长时间收缩为特征的运动。这种全胃性收缩缓慢而有力,它可使胃内压升高,压迫食靡向幽门部移动,并可使食物紧贴胃壁,容易与胃液混合。
二是蠕动,胃壁肌肉紧张与收缩交替运动,这种运动开始于贲门部的细而浅的小波,呈波浪式向幽门部推进,到胃中部时才明显至幽门部,蠕动变强而有力。
作用:使食团与胃液充分混合至半流体食靡,另一方面使胃内容物向幽门部移行。
2、胃运动的调节
1)胃的基础电节律
起源于胃大弯上部,沿纵行肌向幽门方向传播,每分钟3次,其传播速度由大弯向幽门逐渐加快。胃大弯上部平滑肌去极化的频率较胃的其它部分为高,故把它叫胃运动的起步点,在基础电节律基础上产生的动作电位常伴有胃蠕动。
2)神经调节迷走神经对胃运动具有兴奋作用,交感神经(内脏神经)具有抑制作用。交感神经能减慢胃基础电节律的频率和传导速度,抑制胃运动,还有反射性调节或受大脑皮质的影响。
胃运动的反射性调节不仅有非条件反射,也有条件反射。如动物看到食物的外形或嗅到食物的气味,均会引起胃运动加强。
3)体液调节许多胃肠道激素都能影响胃收缩和电活动。胃泌素可使胃的基础电节律及动作电位的频率增加、胃运动加强。促胰液素和抑胃肽可使胃运动减弱。
3、胃的排空
食糜由胃排入十二指肠的过程称为胃的排空(gastric emptying)。草食动物胃的排空比肉食动物慢,胃排空的速度取决于饲料的性质和动物的状况。水、粥、粗硬,动物惊慌疲劳时,排空抑制。食靡由胃分批的进入小肠,其动力取决于幽门两边(胃内和十二指肠内)的压力差,胃的消化性运动提高胃内压力,促进排空。
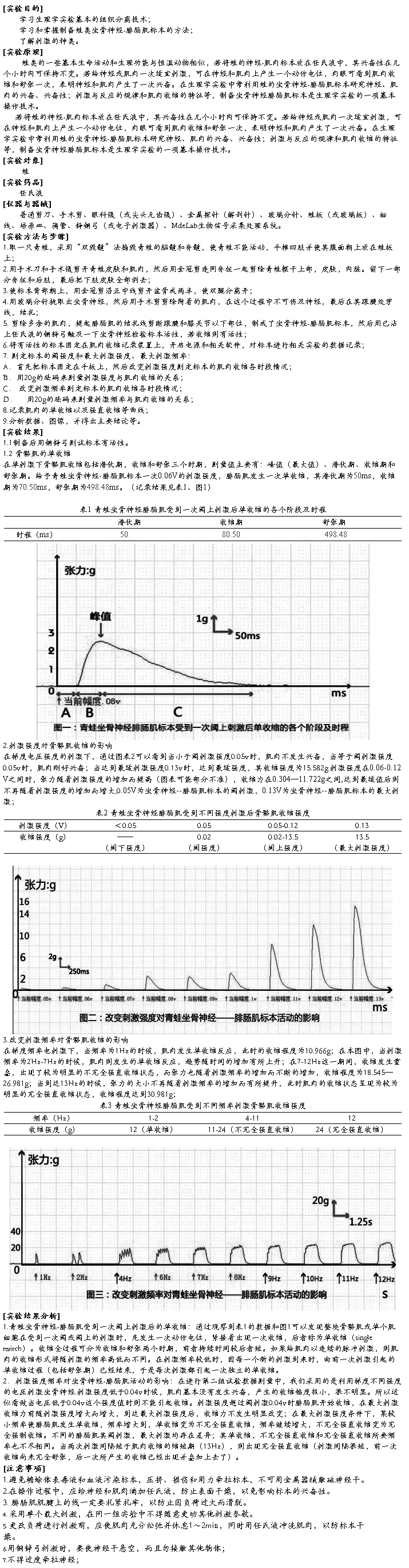
排空的调节:
食靡进入十二指肠抑制胃运动,这是由于食靡入肠后,其中的酸、脂肪以及渗透压过高过低等均可刺激十二指肠壁的感受器,反射的引起胃运动减弱,胃排空减慢,这种反射叫做胃-肠反射,其反射的冲动通过迷走神经、壁内神经从等途径传至胃壁肌,反馈性地抑制胃运动(神经调节)。另外,食靡还可刺激小肠粘膜释放促胰液素、促胰酶素、抑胃多肽等,经体液途径抑制胃运动、延缓胃的排空。
4、呕吐
呕吐(vomiting)是一种保护性反应,排出有害物质。
呕吐动作:一次不随意地深吸气之后,声门关闭,而后胃和食管下端舒张,小肠和幽门部发生逆蠕动,隔肌和腹肌强力收缩,压迫胃内容物从食管和口腔排出。
呕吐是复杂的反射动作,舌根、咽、食管、胃肠心脏、子宫、胃、膀胱、半规管等地感受器受到刺激均可引起呕吐。
传入延髓网状结构地呕吐中枢→迷走N,交感N,隔N,脊N→胃、小肠、隔肌、腹肌→呕吐
第四节复胃内的消化
瘤胃
网胃 前胃:无腺体分布
复胃 瓣胃
皱胃 第四胃(又称真胃):有腺体分布。
一、瘤胃和网胃内消化
食团→前背囊→精料进入网胃,稿杆浮于瘤胃内容物上层,瘤胃下层为液状物,中层多为粗料,上层贮积气体。
瘤胃和网胃是个发酵地大缸,饲料内可消化的干物质70-85%和50%粗纤维在瘤胃内消化,可产生挥发性脂肪酸(VFA)、CO2、NH3以及合成Pr和VB族,因此,瘤胃微生物具有十分重要的作用。
(一)瘤胃内微生物及其生存条件
瘤胃可看作是一个供嫌气性微生物高效率繁殖的发酵罐。它具有微生物活动及繁殖的良好条件,表现在:
1.食物和水分相对稳定的进入瘤胃,供给微生物繁殖所需的营养物质。
2.节律性瘤胃运动将内容物搅拌混和,并使未消化的食物残渣和微生物均匀地排入后段消化道。
3.瘤胃内容物的渗透压与血液相近,并维持相对恒定。
4.适宜的温度。由于微生物的发酵活动,使瘤胃内的温度高达39~41℃。
5.pH值变动于5.5~7.5之间。饲料发酵产生的大量挥发性脂肪酸不断地被吸收入血液,或被随唾液进入的大量碳酸氢盐所中和,以及瘤胃食糜经常地排入后段消化道,使pH值维持在一定范围之内。
6.内容物高度乏氧。瘤胃背囊的气体主要为二氧化碳、甲烷及少量氮、氢等气体,随食物进入的一些氧气,很快会被微生物繁殖所利用。
(二)瘤胃微生物的种类及作用
瘤胃的微生物除了包括细菌和原虫两大类外,还能见到其它类型,如酵母样微生物和 吞噬体。
1、典型的瘤胃微生物应具备如下条件:
(1)必须厌氧性生活。
(2)必须能产生瘤胃内所见的终产物类型
(3)瘤胃内数量必须不少于100万/克内容物
2、纤毛虫
纤毛虫分为全毛和贫毛两类。前者全身被覆纤毛,后者纤毛集中成簇,只分布在一定部位。
作用:(1)发酵糖类,产生乙酸、丁酸、乳酸、CO2、H2等。
(2)分解淀粉。
(3)水解脂类、氢化不饱和脂肪酸。
(4)降解蛋白质及吞噬细菌。
影响纤毛虫种群的因素:
(1)反刍动物种别(2)日粮(3)饲喂次数
(4)饲料加工(5)生理状况(6)昼夜周期性变化
3.瘤胃细菌
种类:纤维素消化菌、半纤维素消化菌、淀粉分解菌,利用糖类细菌、利用酸菌、蛋白分解菌、产氨菌。
作用:(1)消化纤维素(2)分解糖、脂肪、蛋白质(3)合成蛋白质和维生素(B族)。
影响因素:(1)饲料性质(2)饲喂制度(3)饲喂后时间(4)个体差异
(5)纤毛虫存在与否
(三)瘤胃内的消化代谢过程
1.糖类的发酵
可溶性糖发酵最快,淀粉次之,纤维素和半纤维素较缓慢。
纤维素是反刍动物饲料中的主要糖类,其中大约有40~45%在瘤胃内经细菌和纤毛虫的协同和相继作用,首先分解生成纤维二糖,继续分解成葡萄糖,然后经乳酸和丙酮酸阶段而生成VFA、甲烷和二氧化碳。其它糖类通过不同细菌和纤毛虫的发酵,最终产物也大都是VFA、CH4和CO2。VFA主要是乙酸、丙酸和丁酸。
瘤胃微生物在发酵糖类的同时,还能够把分解出来的单糖和双糖转化成自身的糖原,储存于细胞内,当它们随食糜进入皱胃和小肠后,微生物糖原可以被动物所消化利用,成为反刍动物机体的葡萄糖来源之一。
2.蛋白质的消化和代谢
反刍动物能同时利用饲料中的蛋白氮和非蛋白氮,构成微生物蛋白质供机体利用。
进入瘤胃的饲料蛋白,一般约有30~50%未被瘤胃微生物分解而排入后段消化道,其余则在瘤胃内被微生物蛋白酶水解为游离氨基酸和肽类,随后被微生物脱氨基酶分解,生成氨、二氧化碳和短链脂肪酸。因此,瘤胃液中的游离氨基酸很少。畜牧生产中将饲料蛋白质用甲醛溶液或加热法进行预处理后饲喂牛、羊,可以保护蛋白质,避免瘤胃微生物的分解,从而提高蛋白质日粮的利用率。
氨基酸分解所产生的氨,以及微生物分解饲料中的非蛋白含氮物如尿素、铵盐、酰胺等所产生的氨,除了一部分被细菌用作氮源,合成菌体蛋白;另一部分被瘤胃上皮迅速吸收,并在肝脏中经鸟氨酸循环生成尿素。一部分尿素能通过唾液分泌或直接通过瘤胃上皮进入瘤胃,并被细菌分泌的尿素酶重新分解为二氧化碳和氨,可被瘤胃微生物再利用,通常将这一循环过程称为尿素再循环。部分尿素随尿排出体外。尿素再循环对于提高饲料中含氮化合物的利用率具有重要意义,尤其在低蛋白日粮的条件下,反刍动物依靠尿素再循环可以节约氮的消耗,保证瘤胃内氮的浓度,利于瘤胃微生物菌体蛋白的合成,同时使尿中尿素的排出量降到最低水平。
瘤胃微生物合成蛋白质所需的能量和碳源来源于糖、VFA和二氧化碳。
3.脂肪的消化和代谢饲料中的脂肪大部分被瘤胃微生物彻底水解,生成甘油和脂肪酸等物质。其中甘油发酵生成丙酸,少量被转化成琥珀酸和乳酸;来源于甘油三脂的不饱和脂肪酸经加水氢化,转变成饱和脂肪酸。因此反刍动物的体脂和乳脂所含的饱和脂肪酸比单胃动物要高的多。如单胃动物体脂中饱和脂肪酸占36%,而反刍动物则高达55~62%。
细菌还能合成少量特殊的长链或短链的奇数碳脂肪酸、支链脂肪酸,以及脂肪酸的各种反式异构体和立体异构体。
瘤胃微生物的脂肪酸合成受饲料成分的制约,当饲料中脂肪含量少时,合成作用增强;反之,当饲料脂肪含量高时,会降低脂肪酸的合成。瘤胃微生物不能贮存甘油三脂,脂肪酸主要是以膜磷脂或游离脂肪酸形式存在。
4.维生素的合成瘤胃微生物能合成多种B族维生素。其中硫胺素绝大部分存在于瘤胃液中,40%以上的生物素、泛酸和吡哆醇也存在于瘤胃液中,能被瘤胃吸收。叶酸、核黄素、尼克酸和维生素B12等大都存在于微生物体内,瘤胃只能微量吸收。此外瘤胃微生物还能合成维生素K。
幼年反刍动物,由于瘤胃发育不完善,微生物区系不健全,有可能患B族维生素缺乏症;在成年反刍动物,当日粮中钴缺乏时,瘤胃微生物不能合成足够的维生素B12,于是出现食欲抑制,幼畜生长不良等症状。
5.气体的产生在瘤胃微生物强烈发酵的过程中,不断产生大量气体。牛一昼夜产生气体600~1300L,主要是二氧化碳和甲烷,还有少量的氮和微量的氢、氧和硫化氢,其中二氧化碳占50~70%,甲烷占30~40%。气体的产量和组成,随饲料种类、饲喂时间的不同而有显著的差异。
犊牛出生后的几个月内,瘤胃内的气体以甲烷为主。随着日粮中纤维素的增加,二氧化碳的量也增加。到六月龄时,达到成年牛水平。正常动物瘤胃内二氧化碳量比甲烷多,但饥饿或气胀时,则甲烷量显著超过二氧化碳量。
二氧化碳主要是由糖类发酵和氨基酸脱羧产生的,小部分是由唾液内的碳酸氢盐中和脂肪酸时产生的,或脂肪酸吸收时透过瘤胃上皮交换的结果。甲烷是瘤胃内发酵的主要终产物,由二氧化碳还原或甲酸分解所产生。
瘤胃中的气体,约1/4通过瘤胃壁吸收入血后经肺排出;一部分为瘤胃微生物所利用;一小部分随饲料残渣经胃肠道排出;但大部分是靠嗳气(eructation)排出。
(四)、前胃运动及其调节
成年反刍动物的前胃能自发地产生周期性运动,其各部分的运动,在神经和体液因素的调控下,密切联系、相互配合、协调运动。
(一)网瘤胃的运动
整个前胃运动从网胃两相收缩开始。第一相收缩程度较弱,只收缩一半,然后舒张(牛)或不完全舒张(羊),此收缩作用使漂浮在网胃上部的粗糙饲料压向瘤胃。第二相收缩十分强烈,其内腔几乎消失。此时网胃如有铁钉等异物,易造成创伤性网胃炎或网胃心包炎。网胃的这种两相收缩大约每30~60秒钟重复一次。反刍时,在两相收缩之前还出现一次额外的附加收缩,使胃内食物逆呕回口腔。网胃收缩的作用是:(1)驱使一部分液体食糜流进瘤胃前庭。(2)驱使比重轻的食糜流进瘤胃背囊。(3)控制部分液状食糜从网瓣口进入瓣胃。(4)促使前庭内的液状食糜逆流而发生逆呕。
当网胃的第二相收缩至高峰时,瘤胃开始收缩。瘤胃的收缩先由前庭开始,沿背囊依次向后背盲囊传播,然后转入后腹盲囊,由后向前传播,最后终止于瘤胃前部。这种起源于网胃两相收缩的收缩运动,称为瘤胃的原发性收缩,这时所描记的收缩波形称为A波。在原发性收缩的同时,食糜也在瘤胃内顺着收缩的次序和方向移动和混合。在A波收缩之后,有时瘤胃还可发生一次独立收缩,这种与网胃的两相收缩无关的独立收缩,称为瘤胃的继发性收缩(或称B波收缩)。B波是由瘤胃本身产生的,收缩波通常开始于腹盲囊或同时开始于腹盲囊和背盲囊,行进到后背囊和前囊,最后到达主腹囊。在瘤胃出现继发性收缩时,动物往往发生嗳气。“B”波频率在采食时为“A”波的2/3,而在静息时大约为“A”波的1/2。
(二)瓣胃运动
瓣胃运动是与瘤胃运动互相协调的,网胃收缩时,网瓣胃口开放,特别是在网胃第二相收缩时,网瓣胃口开放,此时一部分食糜由网胃快速流入瓣胃。食糜进入瓣胃后,瓣胃沟首先收缩,使其中的液态食糜由瓣胃移入皱胃,而固态食糜则被挤进瓣胃的叶片之间,在瓣胃收缩时可进一步对其进行机械磨碎作用。瓣胃沟的收缩通常与瘤胃背囊收缩同步,恰好在网胃两相收缩的间歇期。紧接着,瓣胃体也发生1~2次收缩,食糜通过开放的瓣皱孔进入皱胃。瓣胃推移食糜的速度受瘤—网胃和皱胃内食糜容量的控制。当网瘤胃内食糜容量增多或皱胃内食糜容量减少时,瓣胃推移食糜的速度加快。有时,当瓣胃体收缩时,瓣皱孔关闭而网瓣孔开放,部分瓣胃内食糜被推回网胃。其功能可能是清除瓣胃沟内的较大颗粒状食糜。
瓣胃具有吸收功能,然而此功能的意义尚有待深入研究。重要的是在食糜被推送进皱胃之前,食糜中残存的VFA和碳酸氢盐已被吸收,避免了对皱胃的不良影响,保证皱胃消化功能的正常进行。
(三)前胃运动的调节
反刍动物的胃运动也象单胃动物一样,具有自动节律性。在正常情况下,这种节律性受神经系统的调节,其基本中枢位于延髓,高级中枢位于大脑皮层,中枢的传出冲动经迷走神经和交感神经传到前胃,支配其节律性活动。各种体液因素也参与胃运动的调节。
切断两侧迷走神经后,食糜不能由瘤胃和网胃进入瓣胃和皱胃,前胃各部出现彼此没有任何连贯性和协调性的收缩。但皱胃运动仍可维持,并可有效地排空,这可能与壁内神经丛的活动有关。如果切断迷走神经的胃支,再刺激其外周端,可引起前胃各部的有力收缩。刺激交感神经的外周端,可抑制前胃各部的收缩。
刺激口腔感受器(如咀嚼)或前胃的张力和化学感受器,都能反射性的引起前胃运动加速加强。刺激网胃的感受器还可以引起逆呕和反刍。消化管各部的状态对前胃运动也有影响。例如,皱胃充满时,瓣胃运动减弱减慢;瓣胃充满时,瘤胃和网胃运动减弱;刺激十二指肠感受器常引起前胃运动的抑制。
前胃运动也受大脑皮层的控制。当某些外来的刺激如噪声、生人出现时,会通过视、听等感觉通路反射性引起瘤胃运动的减弱和反刍停止。而不受干扰、处于安静状态的反刍动物,其副交感神经较为活跃,胃的运动也更强。
胃肠道激素如促胰液素、胆囊收缩素等对瘤胃运动有抑制性作用;胃泌素对瘤胃运动有兴奋作用。
(五)反刍
反刍(rumination)是指反刍动物将没有充分咀嚼而咽入瘤胃内的饲料经浸泡软化和一定时间的发酵后,在休息时返回口腔仔细咀嚼的特殊消化活动。反刍分为四个阶段:逆呕(regurgitation)、再咀嚼(remastication)、再混入唾液(reinsalivation)和再吞咽(redeglutation)。反刍的生理意义在于动物可以在短时间内尽快地摄取大量食物,贮存于瘤胃中,然后在休息时将食物逆呕回口腔,充分咀嚼。是反刍动物在进化中逐渐发展起来的一种生物学适应,借以避免在采食时受到各种肉食动物的侵袭。其功能是将饲料嚼细并混入大量唾液,以便更好地消化。
在个体发育的过程中,反刍动作的出现是与摄取粗饲料相联系的。犊牛大约在出生后的20~30周龄开始选食青草,瘤胃也开始具备发酵的条件,这时动物开始出现反刍。成年牛喂饲干草时,每天反刍时间可长达8小时;如喂饲切碎的干草或精饲料,反刍时间明显缩短。反刍动物一般在采食后0.5~1小时开始反刍,每次反刍通常可持续40~50分钟。然后间歇一段时间再开始下次反刍。成年牛每昼夜大约进行6~8次反刍,幼畜次数更多。反刍易受环境的影响,惊恐、疼痛等因素可干扰反刍,使反刍抑制;发情期、热性病和消化异常时,反刍减少。所以,正常的反刍是反刍动物健康的标志之一。
反刍时,网胃在两次收缩之前产生一次附加收缩,使一部分胃内容物上升到贲门口。然后贲门扩张,动物关闭声门吸气,胸内负压加大,食管内压下降4.00~5.33kPa,胃内容物进入食管,由食管的逆蠕动将食物以大约1m/s的速度返回口腔,这一过程叫逆呕。逆呕是一个复杂的反射活动,是由粗糙食物刺激了网胃、瘤胃前庭和网胃沟(食管沟)粘膜感受器经传入神经(迷走神经)传到延髓的逆呕中枢,中枢的兴奋沿传出神经(主要是迷走神经)和与逆呕有关的膈神经和肋间神经传到网胃壁、网胃沟、食管、呼吸肌以及与咀嚼和吞咽有关的各肌群,引起逆呕动作,开始反刍。当网胃和瘤胃中的食糜经过反刍和发酵变成细碎颗粒时,一方面对瘤胃前庭和网胃等的刺激减弱,另一方面细碎的小颗粒食糜转入瓣胃和皱胃,刺激其感受器,反射性抑制网胃收缩,逆呕停止,进入反刍的间歇期。在间歇期内,瓣胃和皱胃的食糜相继进入小肠,解除了对瓣胃和皱胃的刺激,因而对网胃收缩的抑制作用渐渐解除。当网胃、瘤胃前庭和网胃沟(食管沟)粘膜感受器再次受到饲料刺激时,逆呕重新开始,发生新一轮的反刍。
(六)嗳气
瘤胃中气体部分通过食管向外排出的过程,称为嗳气(eructation)。牛每小时约嗳气17~20次。嗳气的次数决定于气体产生的速度。正常情况下瘤胃中所产生的气体和通过嗳气等所排出的气体之间维持相对平衡。如产生的气体过多,不能及时排出,可形成瘤胃急性臌气。气体正常时积聚于瘤胃背囊的顶部,而背囊的液面高于食管开口。只有背囊前肌柱和后肌柱同步收缩时,气体才能向前和向腹面流动而进入食管口。因此,嗳气一般都在背囊发生继发性收缩时出现。牛的嗳气频率大约是0.6次/分。
嗳气是一种反射动作,它是由于瘤胃内气体增多,对瘤胃背囊壁的压力增大,兴奋了瘤胃背囊和贲门括约肌处的牵张感受器,经迷走神经中的传入纤维,传到延髓嗳气中枢,中枢的兴奋通过迷走神经传出引起背囊收缩,收缩波由后向前推进(B波)压迫气体进入瘤胃前庭,同时前肌柱和网瘤胃褶收缩,阻挡液状食糜前涌;贲门区的液面下降,贲门括约肌舒张,气体向前和向腹面流动而进入食管。当气体充满食管时,贲门括约肌关闭,咽—食管括约肌舒张,气体被迫由食管进入鼻咽腔。此时,鼻咽关闭,声门开放,少量气体从口腔逸出(占0.5~33%),其余的气体经气管进入肺脏,并随呼吸呼出体外。
各种麻醉剂和胆碱能阻断剂都能抑制瘤胃运动。饱食后应用这些药物必须注意移除瘤胃内积聚的气体。当日粮中谷类比例过多或采食大量嫩豆科植物时,可能会发生急性瘤胃臌涨。因为这些食物能较快地进入皱胃和小肠,使其中压力升高,反射性抑制瘤胃运动,使嗳气发生障碍。同时,由于瘤胃内气体不断增多,而渗入瘤胃内容物,形成微小泡沫,无法再行排出。因此治疗时可使用大蒜制止发酵,同时利用植物油或其它不被吸收的表面活性剂破坏瘤胃内形成的泡沫,来预防和治疗瘤胃胀气。
(七)食管沟(网胃沟)反射
网胃沟起始于贲门,向下延伸至网—瓣胃间孔。网胃沟实质上是食管的延续,收缩时呈管状,起着将乳汁或其它液体自食管输往瓣胃沟和皱胃的通道作用。
食管沟反射与吞咽动作是同时发生的,感受器分布在唇、舌、口腔和咽部的粘膜上,传入神经为舌咽神经、舌下神经和三叉神经的咽支,反射的中枢位于延髓内,与吸吮中枢紧密相关。传出神经为迷走神经。若切断两侧迷走神经,网胃沟闭合反射就会消失。幼畜哺乳时,吸吮动作可反射性的引起食管沟的两唇闭合成管状,形成将乳汁通向皱胃的直接通道。这种反射活动在断奶后伴随年龄的增长逐渐减弱以至消失。但如果一直连续喂奶,则到成年时仍可保持幼年时的机能状态。
食管沟有两种收缩方式,一种是闭合不完全的收缩,两唇仅是缩短变硬,两侧相对形成通道,约有30~40%的液体流经食管沟进入皱胃;另一种是闭合完全的收缩,即两唇内翻,形成密闭的管状,摄入的流质食物有75~90%由此流入皱胃。
食管沟反射受多方面因素的影响。(1)动物的摄乳方式。如当犊牛用桶饮乳时,食管沟闭合不完全,乳汁容易进入网胃和瘤胃。由于网瘤胃发育不完善,漏入的乳汁不能顺利排出,时间长后易发生酸败而引起腹泻。当用人工哺乳器慢慢吸吮时,食管沟闭合完全。(2)某些无机盐类有刺激食管沟使其闭合的作用。Cu2+和 Na+对羊作用明显,如CuSO4、NaHCO3等;对牛来说,Na+比Cu2+更为有效,如NaCl、NaHCO3等。在兽医实践中,往往借助上述溶液对食管沟反射的刺激作用,先给予上述溶液,再投药能使药物直接经食管沟进入皱胃发挥作用。
三、皱胃内消化
皱胃的功能与单胃动物相类似。
- 分泌特点:有腺部分,分胃底和幽门两部分。
- 皱胃液连续分泌。
- 胃底腺分泌多,幽门腺分泌少。
- 胃液高度酸性(VFA),可分解微生物蛋白。
- 与单胃动物比较,皱胃液盐酸浓度较低,凝乳酶含量高。
- 皱胃分泌胃液也受神经和体液的调节,刺激交感N,分泌增加;PH降低时,可抑制胃泌素的释放,酸分泌减少。故皱胃PH=2-2.5左右。
- 皱胃运动:在绵羊上研究表明,胃体部一般处于静止状态,只在幽门窦运动明显。
第五节小肠内的消化
食糜由胃进入十二指肠后,受到胰液、胆汁和小肠液的化学作用及小肠运动的机械作用,大部分营养物质在此被吸收和利用。
一、胰液
(一)、胰液分泌收集方法:通常采用插入胰导管的方法、接桥法(体外吻合现在应用T形管方法)。
(二)、胰液的分泌
内分泌部
胰腺
外分泌部:腺泡分泌胰液,胰液是无色透明的碱性液体,PH7.2-8.4。有水分、电解质(高浓度的碳酸氢盐和氯化物)和有机物构成。有机物主要为蛋白质构成的消化酶。
- 胰蛋白分解酶:主要包括胰蛋白酶、靡蛋白酶(A、B)和羧基肽酶等。作用:水解蛋白质为肽及氨基酸等。
- 胰脂肪酶:
- 胰淀粉酶
- 其它
(三)、胰液分泌的调节
1.神经调节
支配胰腺的神经为交感N和副交感N。
刺激交感N可引起少量胰液分泌。迷走神经含有兴奋和抑制胰液分泌的两类纤维。
2.体液调节调节胰液分泌的体液因素主要有:
(1)促胰液素酸性食糜可促进小肠粘膜“S”细胞释放促胰液素。小肠上段粘膜含促胰液素较多,距幽门越远,含量越少。使其释放的最强刺激因素是盐酸,其次为蛋白质分解产物和脂酸钠,糖类几乎没有作用。促胰液素释放与迷走神经兴奋无关,切除小肠的外来神经,盐酸在小肠内仍能引起胰液分泌,说明促胰液素的释放不依赖于肠管外来神经。
促胰液素主要作用于胰腺小导管的上皮细胞,使其分泌大量的水分和碳酸氢盐,因而使胰液的分泌大为增加,但酶的含量却很低。
(2)胆囊收缩素是小肠的“I”细胞释放的一种33肽激素,引起胆囊收缩素释放的因素(由强至弱)为:蛋白质分解产物、脂酸钠、盐酸、脂肪、糖类没有作用。
胆囊收缩素的主要作用为:促进胰液中各种酶的分泌,因而也称促胰酶素;促进胆囊强烈收缩,排出胆汁;对胰腺组织具有营养作用,促进胰组织蛋白质和核糖核酸的合成。
(3)胃泌素由胃窦粘膜和十二指肠粘膜“G”细胞分泌的胃泌素,对胰液中水和碳酸氢盐分泌的作用较弱,而对胰酶分泌的作用较强。
此外小肠分泌的血管活性肠肽具有与促胰液素相似的作用。
近年来的资料表明,促胰液素和胆囊收缩素对胰液分泌的作用是通过不同的机制实现的,前者以cAMP为第二信使,后者则通过磷脂酰肌醇系统,在Ca2+的介导下起作用的。促胰液素和胆囊收缩素之间还具有协同作用,迷走神经对促胰液素的分泌也有加强作用。当诸因素同时作用于胰腺时,将引起胰液更强烈分泌。
二、胆汁
胆汁(bile)是由肝细胞周期性连续分泌的。它不仅是消化液,对食物中的脂肪消化和吸收起着重要作用,而且某些代谢终产物也通过胆汁经肠道排出体外。 马、驴、鹿、骆驼、象、长颈鹿、大鼠、鸽等没有胆囊,其功能在一定程度上由粗大的胆管所取代。由于Oddi括约肌明显地失去功能,胆汁几乎连续地排入十二指肠。
(一)胆汁的性质、成分和作用
胆汁是一种具有苦味的粘滞性有色液体。
胆汁的主要成分有胆汁酸、胆盐、胆色素、胆固醇、粘蛋白、卵磷脂和其他磷脂、脂肪酸和各种电解质,但没有消化酶。
胆盐是肝细胞分泌的胆汁酸与甘氨酸或牛磺酸结合形成的钠盐或钾盐,它是胆汁参与消化和吸收的主要成分。胆汁酸有游离胆汁酸和结合胆汁酸两大类,以后者为主。胆色素是血红蛋白的分解产物,包括胆红素及其氧化产物胆绿素。
胆汁的生理作用主要是胆盐或胆汁酸的作用,主要有:(1)降低脂肪的表面张力,使脂肪乳化成微滴,分散于肠腔,增加了与胰脂肪酶的接触面积,促进脂肪的分解。(2)胆盐因其分子结构特点,当达到一定浓度时,可与甘油一酯和脂肪酸结合成水溶性的混合微胶粒,使脂肪分解产物以及脂溶性维生素(A、D、E和K)能到达肠粘膜的表面,促进其吸收。(3)胆盐本身是促进胆汁分泌的重要体液因素。胆汁中的胆盐和胆汁酸进入小肠后,绝大部分(约90%以上)可以在回肠末端被主动吸收,经由门静脉返回肝脏,然后再分泌到胆汁中去,这一过程称为胆盐的肠肝循环(enterohepatic circulation),见图5-19。(4)增强脂肪酶的活性,起激动剂作用。(5)胆盐可刺激小肠运动。
(二)胆汁分泌的调节
1.神经调节采食动作或饲料对胃和小肠的刺激,可反射性的引起肝胆汁分泌的少量增加,并使胆囊收缩也轻度加强。反射的传出途径是迷走神经,并通过其末梢释放乙酰胆碱直接作用于肝细胞和胆囊平滑肌细胞,也可通过迷走神经—胃泌素途径间接引起肝胆汁的分泌和胆囊收缩。交感神经兴奋可引起Oddi括约肌收缩和胆囊平滑肌的舒张,抑制胆汁的排出。
2.体液调节
(1)促胰液素促胰液素的主要作用是刺激胰液分泌,同时还有刺激肝胆汁分泌的作用。它主要作用于胆管系统而不是肝细胞,其结果是引起胆汁分泌的量和碳酸氢盐含量增加,而胆盐的含量并不增加。
(2)胆囊收缩素在蛋白质分解产物、盐酸和脂肪等物质作用下,小肠粘膜内的I细胞可释放胆囊收缩素,引起胆囊平滑肌的强烈收缩和Oddi括约肌的紧张性降低,从而引起胆汁的大量排放。胆囊收缩素也能刺激胆管上皮细胞,使胆汁流量和碳酸氢盐的分泌增加,但作用较弱。
(3)胃泌素胃泌素对肝胆汁的分泌和胆囊平滑肌的收缩均有一定的刺激作用,它既可以直接作用于肝细胞和胆囊,也可以通过先引起胃酸分泌,之后由胃酸作用于十二指肠,引起促胰液素的分泌而间接起作用。
(4)胆盐胆盐可通过肠肝循环回到肝脏,刺激肝胆汁的分泌。
此外,生长抑素、P物质、促甲状腺激素释放激素等脑肠肽,具有抑制胆汁分泌的作用。
三、小肠液
小肠内有两种腺体:十二指肠腺和肠腺。十二指肠腺又称为勃氏腺(Brunner,s gland),分布在十二指肠的粘膜下层中,分泌碱性液体,内含粘蛋白,因而粘稠度很高。其主要机能为保护十二指肠的粘膜上皮不受胃酸的侵蚀。肠腺又称为李氏腺(Lieberkühn crypt),分布于全部小肠的粘膜层内,其分泌液构成小肠液的主要成分。
(一)小肠液的性质、成分和作用
小肠液是一种弱碱性液体,pH约为8.2~8.7,其中的无机物来自血浆,渗透压与血浆相等。小肠液中的有机物主要是粘液、多种消化酶和大量脱落的肠粘膜上皮细胞。大量的小肠液可以稀释消化产物,利于消化和吸收。小肠液分泌后又很快被小肠绒毛吸收,小肠液的这种循环交流,为小肠营养物质的吸收提供了媒介。
近年来认为,真正由小肠腺分泌的酶只有肠致活酶一种小肠液,它能激活胰液中的胰蛋白酶原,使之变为有活性的胰蛋白酶,从而有利于蛋白质的消化。小肠液中大部分的酶都来源于小肠粘膜的脱落上皮,在细胞崩解后释出。肠液中的酶有(1)肠肽酶,主要是氨基肽酶,它可从肽链的氨基端进一步水解多肽。(2)肠脂肪酶,能补充胰脂肪酶对脂肪水解的不足。(3)二糖酶,主要有蔗糖酶、麦芽糖酶和乳糖酶,分别水解相应的二糖为单糖。虽然这些来源于肠粘膜上皮的酶可以将食糜中的营养物质进一步水解为小分子状态,但对小肠消化并不起主要作用。
(二)小肠液分泌的调节
小肠液的分泌是持续进行的,但不同条件下,分泌量的变化可以很大。食糜对肠粘膜的直接刺激和化学刺激都可引起小肠液的分泌。小肠粘膜对扩张刺激最为敏感。一般认为,这些刺激是通过肠壁内神经丛的局部反射而引起的,且小肠内食糜的量越多,分泌也越多。刺激迷走神经可引起十二指肠腺的分泌,但对其它部位的肠腺作用不明显,有人认为,只有在切断内脏大神经后,刺激迷走神经才能引起小肠液的分泌。
在胃肠激素中,胃泌素、促胰液素、胆囊收缩素和血管活性肠肽都有刺激小肠液分泌的作用。
四、小肠运动
小肠的运动机能是依靠肠壁的两层平滑肌完成的。肠壁的外层是纵行肌,内层是环行肌。小肠的运动可以分为两个时期:一是发生在进食后的消化期,二是发生在消化道内几乎没有食物的消化间期。
(一)小肠运动的形式
小肠的运动形式包括消化期的紧张性收缩、分节运动和蠕动,以及消化间期的移行性运动复合波。
1.紧张性收缩小肠平滑肌的紧张性是其它运动形式有效进行的基础。当小肠紧张性降低时,肠壁易于扩张,小肠对食糜混合无力,推送缓慢。紧张性高时,食糜在肠腔内的混合和推送过程均加快。
2.分节运动(segmentation)主要由肠壁的环行肌的收缩和舒张所形成。在食糜所在的某一段肠管上,环行肌在许多点同时收缩,把食糜分割成许多节段。随后,原来收缩处舒张,而原来舒张处收缩,使食糜的节段分成两段,而相邻的两半则合拢以形成一个新的节段。如此反复进行,食糜得以不断地分开,又不断地混合(图5-20)。当持续一段时间后,由蠕动把食糜推到下一段肠管,再重新进行分节运动。分节运动的主要生理功能是:(1)使食糜与消化液充分混合,便于进行化学消化。(2)使食糜与肠管紧密接触,有利于吸收。分节运动还能挤压肠壁,有助于血液和淋巴的回流。
分节运动只发生在消化期。小肠各段分节运动的频率不同,小肠上部频率较高,下段较低,呈梯度递减趋势。这种梯度现象是由平滑肌的基本电节律所决定的。
3.蠕动小肠的蠕动可发生在小肠的任何部位,其速度约为0.5~2.0cm/s,近端小肠的蠕动速度大于远端。小肠的蠕动波很弱,通常只能推进很短的一段距离,然后消失。其作用在于使经过分节运动作用的食糜向后推进一步,到达下一肠段,再进行新的分节运动。在小肠还常见到一种进行速度快、传播较远的蠕动,称为蠕动冲。蠕动冲可把食糜从小肠始端一直推送到末端,有时还可推送到大肠。蠕动冲可能是由于进食时吞咽动作或食糜进入十二指肠而引起的。
在十二指肠和回肠末段有时还会出现与蠕动方向相反的蠕动,叫做逆蠕动(antiperistalsis)。食糜可以在两段肠管内来回移动,有利于食糜的充分消化和吸收。
4.移行性运动复合波(migrating motility complex, MMC)这是发生在消化间期的一种强有力的蠕动性收缩,传播很远,有时能传播至整个小肠。它发生于胃或小肠上部,沿肠管向肛门方向传播,在传播途中速度逐渐减慢。当一个波群到达回盲部时,另一波群又在十二指肠发生,其间隔通常为90~120分钟。MMC的生理意义尚不太清楚,一般认为在推送小肠内未消化的食物残渣离开小肠和控制前段肠管内细菌的数量方面起重要作用。MMC的发生和移行受神经和激素的调节。迷走神经兴奋可使周期缩短,消化间期胃动素的分泌与MMC的发生有关。
(二)小肠运动的调节
1.内在神经丛的作用位于纵行肌和环行肌之间的肌间神经丛对小肠运动起主要作用。当机械或化学刺激作用于肠壁感受器时,通过局部反射可引起平滑肌蠕动。切断外来神经,小肠的蠕动仍可进行。
2.外来神经的作用一般来说,副交感神经的兴奋能加强肠的运动,而交感神经兴奋则产生抑制作用。但上述效果还以肠肌当时的状态而定。如肠肌的紧张性高,则无论副交感神经还是交感神经兴奋,都使之抑制;相反,如肠肌的紧张性低,则这两种神经兴奋都有增强其活动的作用。
3.体液因素的作用促进小肠运动的体液因素有:乙酰胆碱、5-羟色胺、胃泌素、胆囊收缩素、胃动素、P物质等。其中P物质、5-羟色胺等作用更强。抑制小肠运动的物质有血管活性肠肽、抑胃肽、内啡肽、促胰液素、肾上腺素、胰高血糖素等。
(三)回盲瓣或回盲括约肌的机能
牛、羊、猪、狗有很发达的回盲瓣,而马有很发达的回盲括约肌。在回盲瓣处,环行肌显著增厚,形成回盲括约肌,平时回盲括约肌保持收缩状态。当食物进入胃时,可通过胃—回肠反射引起回肠蠕动,在蠕动波到达回肠末端最后数厘米时,括约肌舒张。当盲肠粘膜受到机械刺激或扩张刺激时,可通过肠肌局部反射,引起括约肌收缩,阻止回肠内容物向盲肠排放。
回盲瓣或回盲括约肌的主要功能是防止回肠内容物过快地进入盲肠,延长食糜在小肠内停留的时间,保证小肠内容物能更充分地消化和吸收;同时,它还能有效地阻止大肠内有细菌的内容物向回肠倒流而污染小肠。
第六节大肠消化
食糜经小肠消化吸收后,其残余部分逐渐进入大肠。肉食动物的消化吸收过程在小肠内已经基本完成,大肠的基本功能主要是吸收水分和形成粪便。草食动物,特别是单胃草食动物的大肠内容物中还含有较为丰富的营养物质,因而大肠内仍有较为强烈的消化活动,并且在整体消化活动中占相当重要的地位。反刍动物和杂食动物的大肠消化也有一定的作用。
一、大肠液的分泌
大肠液是由大肠粘膜表面的柱状上皮细胞及杯状细胞分泌的富含粘液和碳酸氢盐的液体,其pH值为8.3~8.4。碳酸氢盐的作用在于中和大肠内发酵产生的酸,这在草食和杂食家畜尤为重要。粘蛋白的作用是润滑粪便和保护大肠粘膜不被粗糙的消化残渣所损伤。大肠液的分泌主要是由大肠内容物对肠粘膜的机械刺激所引起。分泌活动是在壁内神经丛的控制下进行的。副交感神经兴奋时,大肠液分泌增强;而交感神经兴奋则分泌减少。目前尚未发现大肠液分泌的体液调节证据。
二、大肠的运动和排粪
大肠运动的特点是少而慢,对刺激的反应也较迟钝。这些特点有利于大肠内微生物的活动和粪便的形成。
(一)盲肠运动
肉食动物只有不发达的盲肠,而草食和杂食动物都有发达的盲肠。各种家畜的盲肠都能进行类似小肠分节运动的节律性收缩,但频率和速度都比小肠低的多。它的生理功能是搅拌和揉捏盲肠内容物,没有推进作用。进食过程常使这种运动加快和增强。盲肠中出现的第二种运动是强烈的推进运动。这种运动以不规则的间隔周期性出现,其作用是把盲肠内容物送入结肠。盲肠的第三种运动是蠕动,它使内容物在盲肠中缓慢地移动,最终送入结肠。猫、狗、牛、羊、马等还可在盲肠出现逆蠕动,以延缓内容物在盲肠中停留的时间。猪一般不发生盲肠逆蠕动。
(二)结肠运动
各种家畜都有发达的结肠。结肠基本电节律的起步点是前结肠或结肠中段的环行肌。由此产生慢波沿结肠分别向两端传播:即前半段向近端传播,后半段向远端传播。结肠中出现的运动形式有:袋状往返运动、分节或多袋推进运动和蠕动。
1.袋状往返运动这是在空腹时最多见的一种运动形式,由环行肌无规律地收缩所引起,使结肠形成多个袋状结构,并使结肠袋中的内容物向两个方向作短距离位移,并不向前推进。在结肠后段,这种运动与形成卵圆形的粪块有关。袋状往返运动除速度缓慢和波及的肠段较长外,性质上类似小肠的分节运动。
2.分节或多袋推进运动这是一个结肠袋或一段结肠收缩,将其内容物推送到下一段的运动。进食后或结肠受到拟胆碱药物刺激时,这种运动增多。其作用是将结肠内容物缓慢地向肛门方向推进。在这种运动之后,随之出现排粪。
3.蠕动、逆蠕动和集团运动结肠蠕动是由稳定的收缩波所组成。蠕动能使内容物以每分钟几厘米的速度向肛门端推进。收缩波远端的肠肌保持舒张状态,并往往充有气体;收缩波近端的肠肌则保持收缩状态,使该段肠段闭合并排空。结肠蠕动与小肠蠕动的不同之处是有强大的向后推进力量。
在大肠还有一种推进速度很快,且推进距离很远的蠕动,称为集团运动(mass peristalsis)。它可能是食糜进入十二指肠,由内在神经丛产生的十二指肠—结肠反射所引起。
牛、羊、马、猫、狗等的结肠前段可发生逆蠕动,而猪则不发生。
(三)排粪动作
排粪动作是由大肠后段的平滑肌和肛门括约肌等一系列肌群参与的复杂反射。排粪时,结肠后段和直肠的肌肉强烈收缩,肛门内外侧括约肌舒张,粪便被排出体外。同时,膈肌和腹肌发生协同收缩,提高腹腔内压,协助粪便排出。引起排粪反射的正常刺激是粪便直接刺激了直肠壁内的压力感受器,冲动经盆神经和腹下神经传至脊髓腰荐段的低级排粪中枢,并由此上传到大脑皮层,引起便意和排粪反射。中枢发出的冲动经盆神经传至大肠后段,引起大肠后段肌肉收缩和肛门内括约肌的舒张;同时,冲动也经相应的脊神经传出,引起腹肌、膈肌收缩,使腹内压增加;肛门外括约肌舒张,使粪便排出体外。
排粪动作受大脑皮层的控制。家畜能随意进行或抑制排粪,也能建立排粪的条件反射。除狗、猫外,家畜能在行进状态下排粪。
三、大肠内的生物学消化
(一)肉食动物大肠内的生物学消化
食糜中没有被小肠消化吸收的蛋白质,可以被大肠中的腐败菌分解生成吲哚、粪臭素(甲基吲哚)、酚、甲酚等有毒物质。这些物质一部分由肠粘膜吸收入血液,在肝脏内经解毒后随尿排出体外;另一部分则随粪便排出。
小肠内没有被消化的脂肪和糖类,在大肠内也经细菌作用,脂肪分解成脂肪酸及甘油,糖类分解为单糖及其它产物,如草酸、甲酸、乙酸、乳酸、丁酸以及二氧化碳、甲烷、氢气等。
(二)草食动物大肠内的消化
马、兔,饲料中的纤维素等多糖物质的消化和吸收,全靠大肠内微生物的作用。
单胃草食动物大肠容积庞大,有大量微生物生长和繁殖,和反刍动物瘤胃相似。
1、使糖类发酵变成VFA或低浓度的乳酸盐、CO2、CH4等。
2、进入大肠的含氨物质(饲料中的Pr,非蛋白含氮物质,脱落的肠上皮)被微生物利用合成大量菌体蛋白。
3、大肠内微生物能合成B族维生素,如VK,并被大肠粘膜吸收。
4、大肠壁还能排泄Ca、Mg、Fe等矿物质。
(三)杂食动物大肠内消化
1、饲料中纤维素和未被小肠消化的可溶性糖在发酵菌作用下,产生有机酸(乳酸、VFA)和气体(CH4、CO2、N2、H2)等。
2、饲料中的Pr、氨基酸和尿素等在腐败菌作用下,产生吲哚、尸胺、粪臭素、氨等。
3、细菌也能合成B族维生素。
(四)大肠运动
与小肠运动相似,也具有蠕动,分节运动及钟摆运动,但速度及强弱等均较弱。
第七节吸收
消化道内营养物质的吸收是指食物的消化产物、无机盐和水等,通过消化道上皮细胞进入血液和淋巴的过程。
一、吸收的部位
胃肠吸收的主要部位在小肠。小肠粘膜表面遍布环状皱褶,上有密集的指状突起-绒毛(intestinal villi)。每一条绒毛的表面被覆一层柱状上皮细胞,称为绒毛细胞。每个绒毛细胞肠腔面的细胞膜向外突出,形成许多微绒毛(microvilli),使小肠的吸收面积增加了数十倍数百倍。
小肠绒毛的内部有丰富的毛细血管网和一根以盲端起始的中心乳糜管,远分布有平滑肌纤维,可进行绒毛的收缩和摆动,促进血液和淋巴的流动。反刍动物的前胃和单胃动物的大肠也是重要的吸收器官,可吸收饲料在前胃和大肠内发酵的产物如VFA、氨等。一般动物的胃以及肉食动物和猪的大肠内只能吸收水分和盐类,对有机营养分的吸收则很有限。
二、吸收的机理
可分为简单扩散、易化扩散、主动转运三种。
1、简单扩散:是一种非耗能过程,它的发生主要取决于物理的驱动力,如:渗透压、流体静力压、溶剂拖曳作用、电化学梯度等。
胸腔内物质通过扩散进入血液的通路有四种:
①通过上皮细胞的脂膜。物质首先脱离水介质,并进入膜向疏水脂区,而后通过脂膜层,最后又脱离脂层重新回到水介质中,因此,这类物质的扩散速度主要决定于它的脂肪性能。
②通过小肠上皮的充水管道:主要是水分和水溶性物质。
③通过细胞间的结合点:主要是小分子的电解质。
④通过细胞挤压而出现的空隙,即吸混作用,主要是一些大分子颗粒。
2、易化扩散:与简单扩散有些类似,也是一种非耗能过程,但由于细胞膜上存在特异的载体,因而扩散速度较快。
3、主动转运:是一种可使物质逆浓度梯度的转运过程,这种过程首先必须借助于膜上的特异性载体。载体是一个Pr或脂蛋白,在蛋白质部分,有与携带的物质分子相结合的特异部位,而脂肪部分,可使结合体保持脂溶性,能通过细胞膜自由扩散。载体系统有特异性,细胞膜上可以有几种不同的载体系统,每一系统只运载某些特定的物质,主动转运必须消耗能量,这主要靠ATP的分解供能:Na-K-ATP酶,离子泵。
三、主要营养物质的吸收
(一)糖的吸收
糖以单糖形式被吸收,主要的单糖是葡萄糖、少量半乳糖、果糖。糖在胃中几乎不吸收,在小肠几乎完全被吸收,吸收后入肝。
吸收机制:单糖的吸收是靠载体系统进行的,这种转运具有高度特异性,它使糖的吸收能逆浓度差进行。
钠泵:实际上就是镶嵌在膜的脂质双分子层中的一种特殊蛋白质,它本身具有ATP酶的活性,可以分解ATP获得能量,并用此能量进行Na+、K+ 的主动转运,因此Na泵就是这种被称为Na-K依赖或ATP酶的蛋白质。
载体蛋白在转运糖时,必须先携带Na+,才能与单糖分子结合形成钠泵复合物,当其透过细胞膜入胞浆后释放出糖,Na+然后通过Na+泵放出Na+并耗能。细胞内糖靠扩散方式透过绒毛内的毛细血管,在各种单糖中,己糖吸收最快,戊糖吸收则慢,在己糖中又以半乳糖和G吸收最快,果糖次之,甘露糖最慢。
(二)脂肪的吸收
 脂肪消化后主要形成甘油、脂肪酸、甘油一酯,另外还有甘油二酯和未消化的甘油三酯、胆酸盐+脂肪的水解产物水溶性复合物多个脂肪微粒。
脂肪消化后主要形成甘油、脂肪酸、甘油一酯,另外还有甘油二酯和未消化的甘油三酯、胆酸盐+脂肪的水解产物水溶性复合物多个脂肪微粒。
有人认为:脂肪微粒通过胞饮作用而吸收,另一些人认为:脂肪微粒靠近刷状緣,产物从微粒中释放出来。
脂肪微粒吸收后,脂肪的分解产物在肠上皮细胞重新合成中性脂肪,并在外面包了一层卵磷脂和蛋白质形成的膜,形成乳糜微粒,靠出胞作用进入细胞间隙,由此进入乳糜管,再汇集于淋巴管内血循。
脂肪的吸收以淋巴为主,甘油、短中链脂肪酸通过血液吸收。
(三)蛋白质的吸收(朊)
朊的吸收部位在小肠,尤其是小肠上皮。
朊在小肠内分解为氨基酸而被吸收,吸收是主动过程:机制和糖相似。中性AA比酸性AA吸收快。目前已确定,有三种AA的特殊转运系统,一是转运中性AA,如蛋AA、亮AA,二是转运碱性AA(双胺),如精AA,赖AA,三是对中性AA中脯AA和羟脯AA的转运。
(四)无机盐的吸收
一般来说,钠、钾、铵盐的吸收很快,钙盐则不易吸收。
1.Na+的吸收存在钠泵,因此未主动重吸收。Na+的主动转运取决于细胞内K+浓度,Na+向膜外与K+向膜内转运是耦联进行的。
2.Fe2+的吸收铁的吸收与机体需要量有关。食物中绝大部分以三价铁形式存在,必须还原为Fe2+才能吸收。VitC能将Fe+++还原为Fe++,促进吸收。胃中的HCl入小肠造成酸性环境,可促Fe++的吸收。Fe在小肠上皮细胞吸收:在肠粘膜细胞内氧化成Fe+++,并以铁蛋白形式存在,当细胞内达到一定量时抑制对铁的吸收,铁由C转送入血的速度根据机体的需要量调节。
3.钙的吸收通过主动转运来吸收。维生素D可促进钙的吸收,钙盐只有在水溶性状态下,且不被肠腔中的其它物质(如草酸盐、磷酸盐等)沉淀时,才能被吸收。肠内容物的酸度对钙的吸收有重要影响,pH值约为3时,钙呈离子状态,吸收最好。脂肪分解所产生的脂肪酸对钙的吸收有促进作用。
4.负离子:主要是Cl-、HCO3-的吸收造成正电位,可促进负离子的吸收
5.水的吸收胃吸收较小,大部分在小肠和大肠吸收。吸收的动力是渗透,由于各种营养物质的吸收,使上皮细胞的渗透压升高,从而促进水的吸收。
来源:除了来自饮食和饮水外,大量的是来自消化液的水,如牛:24小时通过十二指肠水约100升,其中75升来自消化液,这些水分有90%被消化道吸收,因此在消化不良时,排出大量的水和无机盐。
(五)维生素的吸收
1.脂溶性维生素的吸收脂溶性维生素A、D、E、K和胡萝卜素可溶解于胆盐微胶粒中,与其它脂肪消化产物一起吸收。
2.水溶性维生素的吸收以单纯扩散的方式被吸收。维生素B12必须与胃粘膜壁细胞分泌的“内因子”结合成复合物,在转运到回肠后与粘膜上皮细胞上的受体结合而被吸收。回肠是吸收维生素B12的特异部位。
第七章 能量代谢和体温调节
第一节概述
动物的生存有赖于不断地与外界环境进行物质交换和能量交换,也即为新陈代谢。新陈代谢包括物质代谢和能量代谢,物质代谢过程中伴随的能量释放、转移和利用叫能量代谢。
- 基础代谢与静止能量代谢
基础代谢:在清醒、静卧、空腹和20度左右的环境温度等条件下的能量代谢。
若消耗的能量以每小时每平方米表面积所散发的热量(千卡)来表示,称基础代谢率。
基础状态:机体在清醒、极其安静的状态下,各种生理活动比较恒定,代谢率较恒定,这时机体释放尿等基本生命活动的消耗,这种生理状况叫基础状态。
静止能量代谢:系食物的特殊动力作用,动物采食后,尽管处于安静状态,而所产生的热量比采食前增加,这种由于摄食而发生的代谢率增加静止能量代谢或食物的生热效应。
- 能量的来源与去路
动物体通过摄食、消化吸收过程摄入营养物质,如糖、蛋白、脂肪,这些都包含有能量,它们在体内经过一系列的生物氧化过程,最终产生二氧化碳、水,同时放出能量,放出能量一部分是热能,另一部分是化学能,化学能转移到ATP的高能磷酸键中作为机体各种活动的能源,最终一部分转化为机械能用于做功,一部分转化为热能。
根据能量守恒定律:
能量输入=能量输出+能量贮存
能量输入>能量输出,能量贮存为正数,组成机体的物质增加,体重增加。反之相反,消耗体内物质。
三、能量代谢的测定方法
(一)直接测热法(direct calorimetry)
就是将机体在一定时间内发散出来的总热量搜集起来的并加以测量的方法。
采用热量计。
(二)间接测热法(indirect calorimetry)
本方法是测定机体在一定时间内的耗氧量和二氧化碳排出量来计算机体的产热量。其理论依据是化学中的定比定律,既在一般化学反应中,反应物的量与产物的量之间成一定的比例关系。例如葡萄糖氧化时,反应式为:
C6H1206+602—→6CO2+6H20+热量
无论糖在体外燃烧或是在体内氧化,1 mol的葡萄糖都要消耗6 mol的氧,产生6 mol的水,产生的热量也相等,这就是一种定比关系。测知了一个数据,就可计算相关的量。
(三)氧的热价及呼吸商
(1)食物的热价每克营养物质完全氧化时所释放的热量,叫做该物质的热价(caloric value)。
(2)食物的氧热价 某种营养物质氧化时,消耗1升氧所产生的热量,叫做该物质的氧热价(thermal equivalent of oxygen)。
(3)呼吸商 生理学上把机体在单位时间内CO2排出量与耗氧量的比值称为呼吸商(respiratory quotient ,简称RQ)。通常以容积数值(L)来表示CO2与O2的比值。即:
CO2排出量(L)
呼吸商RQ=──────────────
耗O2量(L)
糖的呼吸商为1.0,脂肪为0.71,蛋白质为0.80。
间接测热法的具体计算步骤如下:
测定机体一段时间内耗O2量和CO2排出量,计算呼吸商。现今已有多种仪器设备,用以测定动物的耗O2量和CO2的排出量。为了便于比较,所测气体量应换算成标准状况下气体容积,然后计算出呼吸商(混合呼吸商)。
测定一段时间内蛋白质、糖和脂肪的产热量,应首先计算蛋白质的产热量,由于尿中含氮物质主要是蛋白质的分解产物,因此可以通过测定一定时间内尿氮量来计算体内被氧化的蛋白质数量。蛋白质平均含氮量为16%,如有1克尿氮产生,则有6.25克蛋白质被氧化分解。求出蛋白质的量乘上蛋白质的生物热价,即得到蛋白质的产热量,再计算糖和脂肪的产热量。为此需计算非蛋白呼吸商,计算方法是从总耗氧量中减去蛋白质氧化时的耗氧量,从CO2的排出量中减去蛋白质分解时CO2产生量,然后计算的呼吸商即为非蛋白质呼吸商,具体计算过程如下:
蛋白质耗氧量=尿氮量×6.25×0.95(每克蛋白质氧化耗氧量)
蛋白质CO2产生量=尿氮量×6.25×0.76(每克蛋白质氧化CO2产生量)
总CO2产生量 ― 蛋白质CO2产生量
非蛋白呼吸商= ───────—──────────────
总耗氧量 ― 蛋白质耗氧量
实际工作中,常采用简化的计算方法,即测定一定时间内机体的耗氧量和CO2排出量,并求出呼吸商(混合呼吸商),然后在非蛋白质呼吸商表中查出该呼吸商的氧热价,这里用非蛋白质呼吸商代替混合呼吸商,将耗氧量乘上氧热价,得出机体在该时间内的产热量。
二、影响能量代谢的几个因素
(一) 食物的特殊动力效应
动物在采食后一段时间内,虽然同样处在安静状态,而其产热量却比采食前有明显增加,这种由于摄食使机体产生“额外热量”的现象,叫做食物的特殊动力效应(specific dynamic effect)。研究发现,若吃的全是蛋白质食物,则额外产生的热量可达30%;以糖和脂肪为主,生热可增加4%~6%,一般混合食物可增加10%左右。
(二) 肌肉活动的影响
肌肉是机体主要的产热器官。据估测动物在安静时,其产热量占全身总产热量的20%,运动或使役时,肌肉产热量可高达总产热量的90%。所以机体任何轻微的活动,均可提高代谢率。
(三) 环境温度的影响
环境温度的明显变化对机体代谢有重大影响,这主要是机体为保持体温稳定对代谢的调节过程。寒冷情况下,产热量增加,主要是寒冷刺激反射性地引起肌肉紧张性增加,甚至发生寒战,产热增多。一般在20~30℃的环境中,代谢率比较稳定。当环境温度上升到30~45℃时,代谢率将逐渐上升,这可能和体内生化反应加快,发汗机能旺盛以及循环、呼吸机能增强等因素有关。
(四)机体本身因素
1.个体大小 动物体格越小,产热越多。这是因为小动物的单位体重比大动物占有较大的体表面积,而体热又是通过体表面积发散的,因而小动物的单位体重散失的热量较多,为了维持体温,就必须多产热。
2.性别在同样条件下,公畜的基础代谢比母畜高,
3.年龄幼年时基础代谢比成年高,成年后则维持在一定的水平且相当平稳。之后,随着年龄增加,代谢率逐渐下降。
4.营养状况营养优良的动物其基础代谢率比不良者高。
第二节体温及其调节
一、动物的体温及其正常变动
体温(body temperature)指的是机体深部的平均温度,是机体在代谢过程中不断产生热能的结果。
来源:体内温度的来源是体内物质代谢过程中所释放出来的热量。对于恒温动物,正是由于机体在代谢过程中不断有热能的释放,才能保持有机体新城代谢的正常进行。
意义:机体的物质代谢过程是以酶促反应为基础的,而酶类必需在适宜的温度条件下才具有较高的活性,体温升高或降低会影响酶的活性。
(一) 动物的正常体温
由于深部温度,特别是血液温度不易测试,在生理学和畜牧兽医实践中,多以直肠温度代表体温。
(二) 体温的生理波动
1.体温的昼夜波动在白天活动的动物,其体温在清晨时最低,午后最高,一天内温差可达1℃左右。
2.年龄新生幼畜代谢旺盛,体温都比成年家畜高。幼畜在出生后的一段时间内容易受外界温度变化的影响而使体温发生波动,说明体温调节能力还比较弱,不能有效地使体温恒定。因此,对幼畜要加强护理和保温。
3.性别雌性动物体温高于雄性。母畜发情时体温升高,排卵时体温下降。
4.肌肉活动肌肉活动时,代谢增强,产热量增加,导致体温上升。
二、 机体的产热和散热过程
正常体温的维持,有赖于机体的产热过程和散热过程的动态平衡。新陈代谢过程中,不断地产生热量,用于维持体温。同时,体内热量又由血液带到体表,通过辐射、传导和对流以及水分蒸发等方式不断地向外界放散,产热量和散热量取得平衡,体温就可维持在一定水平。
(一) 产热过程
机体所有组织器官均处在合成和分解代谢过程中,因而都产生热量。但它们的产热量有所不同,安静时以内脏器官产生热量最多,其中以肝脏代谢最为旺盛,产热量较多。安静时肝、肠、胃产热量约占身体总产热量的50%,肌肉占20%,脑占10%。运动或使役时骨骼肌是产热的主要器官,其产热量可高达总产热量的2/3以上。草食家畜消化道中饲料由于微生物的发酵分解,产生大量热能,是这类动物体热的重要来源。
(二) 散热过程
机体的主要散热器官是皮肤。一部分热量可通过呼吸、排粪和排尿散失。当外界环境温度低于体表温度时,可通过皮肤以辐射、传导、对流等方式进行散热。当环境温度接近或高于皮肤温度时,则只能以蒸发方式散热。所以皮肤是机体热量散失的重要途径,可占全部散热量的75%~85%。散热过多或散热困难都将严重影响体温恒定。
1.辐射(radiation)散热这是体热以红外线的形式传给外界温度较低物体的一种散热方式。辐射散热量取决于皮肤和环境之间的温度差以及机体辐射面积等因素。
2.对流(convection)散热机体通过与体表接触的气体或液体流动来交换和散发热量的方式,称为对流散热。动物体周围经常有一层同体表接触的空气层,其温度又较体温低,则体热可传给这一层空气。热空气向上升,较冷的空气流来填补,体热即可向外界放散。
3.传导(conduction)散热是指机体的热量直接传给同它接触的较冷物体的一种散热方式。机体深部的热量主要由血液流动将其带到皮肤,再由皮肤直接传给和它相接触的物体。传导不是散热的主要形式。
4.蒸发(evaporation)散热水分蒸发是吸热过程,蒸发1g水可带走2.43KJ(0.58kcal)热,所以体表水分蒸发是一种有效散热途径。出汗对调节散热的重要性有明显的畜种差异,马属动物能大量出汗;牛有中等程度的出汗能力;绵羊可以发汗,但热喘呼吸是主要的散热方式;狗几乎全部依靠热喘呼吸散热;而在啮齿动物即不热喘呼吸也不发汗,它们向毛上吐抹唾液或水来蒸发散热。
三、体温恒定的调节
(一) 神经调节
1.温度感受器
外周感受器:在皮肤、某些粘膜及腹腔内脏等处分布有热感受器和冷感受器两种。它们能接受内外环境温度变化的刺激,把这种刺激转变为N冲动,传向体温调节中枢,发生相应变化。
中枢感受N元:在脊髓、延髓、脑干网状结构以及下丘脑等部位存在有对温度变化敏感的神经元,有热敏感神经元和冷敏感神经元。可感受血温的变化,控制调节中枢的兴奋性。
2.体温调节中枢-调节温度的N细胞群
调节体温的基本中枢在下丘脑。
证明:切除大脑皮质及部分皮质下结构后,只要保持丘脑及其以下的神经结构完整,这种动物的体温就能够在冷环境中保持恒定,即仍具有维持体温恒定的能力。如进一步破坏下丘脑,则动物就再不能维持相对恒定的体温,如果将这种动物暴露在较低温度环境中,直肠温度就迅速下降,证明已失去调节体温的能力。这些事实说明,调节体温的基本中枢在下丘脑。过去曾经认为下丘脑前部是散热中枢,而下丘脑后部为产热中枢,两者功能上协调,共同维持体温恒定。
机体的体温调节机构通过对产热和散热过程的调节,可以使体温保持在某一水平,但为什么有些动物的正常体温能平衡在37℃?目前这个问题还没有确切的答案。近年来有人提出调定点(set point)学说加以解释。所谓调定点通常是指工程上或仪器中的恒温调节控制点,把这个装置上的控制旋钮定在某一温度,如37℃,则可使该系统中的温度稳定在37℃。生理学中,温度调定点学说认为,PO/AH中的热敏感神经元可能在体温调节中,起着调定点的作用,它们类似于仪器的恒温调节装置,可控制体温于一定水平。热敏感神经元对温度的感受有一定的阈值,这个阈值就是体温的稳定点。当体内温度超过阈值时(如37℃),热敏感神经元兴奋,发放的冲动频率增加,促使散热活动加强,同时冷敏感神经元放电频率减少,使产热活动受到抑制,结果体温不致上升。当体内温度低于阈值时,发生和上述相反的变化,于是产热增加,如骨骼肌紧张性增加,皮肤血管收缩,结果体温回升。
体温调节中枢的调定点不是固定不变的。当机体处在高温环境时,皮肤温度感受器受到皮温上升的刺激而兴奋,冲动传入中枢,可使调定点下移,此时,即使中枢温度仍在37℃,也会使散热过程加强,但在正常情况下,调定点上下移动的范围很窄。
(二) 体液调节
1.甲状腺激素是由甲状腺分泌的含碘化合物,这种激素能加速细胞内的氧化过程,促进分解代谢,产热量增加。
2.肾上腺素是肾上腺髓质分泌的胺类激素,其主要作用为促进糖和脂肪的分解代谢,促使产热增加。
 3.缓激肽当外界温度升高时,汗腺一方面分泌汗液,增加蒸发散热,另一方面腺C中的激肽原酶被激活,能使组织液中的球蛋白分解,产生激肽作用与局部血管
3.缓激肽当外界温度升高时,汗腺一方面分泌汗液,增加蒸发散热,另一方面腺C中的激肽原酶被激活,能使组织液中的球蛋白分解,产生激肽作用与局部血管
 舒张增加局部血流量加强散热。
舒张增加局部血流量加强散热。
(三) 机体对冷或热的体温调节过程
1、对寒冷的调节过程
寒冷时可使皮肤温度降低,皮肤感受器兴奋,发出冲动沿传入神经上传到下丘脑前区的热敏感神经元,使之抑制。于是出现产热增加,散热减少,来维持提体温的正常活动。生理反应如下:(1)外周血管:收缩、血流量减少,减少散热;(2)汗腺:分泌汗腺减少;(3)肌肉:出现不协调收缩,出现寒颤,产热增加;(4)腺体:肾上腺素,甲状腺分泌增加,产热增加,非寒颤性产热。
2、对炎热的调节过程
当体内外温度升高,尤其是体内温度升高时(如运动),皮肤和内脏的感受器刺激产生冲动沿传入N,传入丘脑下部或当血温升高时引起热敏N元的冲动增加,通过丘脑下部的体温调节中枢,使机体产热减少,增加散热。
生理反应如下:(1)皮肤血管:舒张,血流量增加,机体深部的热量通过血流到皮肤,使皮肤温度升高,以辐射、传导、对流方式散热;(2)汗腺:活动增加,大量分泌汗液;(3)热性喘息:高温环境下,引起呼吸急促,使呼吸道蒸发散热增加。
3、行为调节
寒冷:蜷缩姿势,减少与寒冷接触面积,长期在寒冷中,被毛增加,皮下脂肪增加,使体表的绝热作用增加。
炎热:逃避阳光照射,到阴凉处。
第八章 泌尿生理
排泄是指动物体将体内代谢终产物、多余物质、进入体内的药物或毒物等排出体外的过程。机体的排泄途径有4条:
(1)以气体的形式通过呼吸排出CO2、少量水分和一些挥发性物质;
(2)以汗液的形式排出部分水分、无机盐和尿素等;
(3)通过肠道以粪便的形式排出胆色素、某些无机盐等;
(4)以尿的形式排出体内大量代谢产物。这是最重要的排泄途径,因为通过尿液排出的代谢产物种类多数量大。
第一节 尿的理化性质及组成
尿的组成

第二节尿的生成
一、肾脏的结构特点
(一)肾单位和肾小管
肾单位(nephron)是肾脏的基本结构和功能单位,由肾小体和肾小管组成。
肾小体包括肾小球和肾小囊。
肾小球是一团毛细血管网,其两端分别与入球、出球小动脉相连。肾小球外面的包囊称肾小囊,由肾小管盲端膨大凹陷形成,分内层(脏层)和外层(壁层),两层之间的腔隙称囊腔,与肾小管管腔相通。
肾小管由近球小管 (proximal tubule)、髓袢和远球小管(distal tubule)组成。近球小管包括近曲小管和髓袢降支粗段。髓袢由髓袢降支和髓袢升支组成;前者包括髓袢降支粗段(近球小管的组成部分)和细段,后者为髓袢升支细段和粗段(远球小管的组成部分)。远球小管包括髓袢升支粗段和远曲小管。远曲小管与集合管相连。
(二)皮质肾单位和近髓肾单位
1、皮质肾单位分布于皮质浅表部,特点:肾小球体积小,入球小动脉的口径比出球小动脉粗,出球小动脉再分成毛细血管后几乎全部分布到皮质部分的肾小管周围,髓袢长,可深入到内髓层,有的甚至到达肾乳头部
(三)近球小体
近球小体(肾小球旁器(juxtaglomerular apparatus,JGA)),由颗粒细胞、系膜(间由入球小动脉的近球细胞:远曲小管的致密斑(macula dense)和间质细胞构成。
近球细胞:
致密斑是指位于远曲小管起始部或髓袢升支粗段处邻近入球小动脉一侧的肾小管上皮细胞,此处的上皮细胞变为高柱状,核密集,色浓染,呈现斑纹隆起,称致密斑;它可感受小管液中NaCl含量的变化,并将信息传递给颗粒细胞,调节肾素的分泌。近球小体主要分布在皮质肾单位。
间质细胞
- 尿的生成过程
尿的生成包括肾小球的滤过作用、肾小管及集合管的重吸收和分泌作用。
(一)肾小球的滤过作用
滤过作用:血液流经肾小球时,从血浆中滤出一些基本不含蛋白质的滤液,这一过程称滤过。
原尿:血液流经肾小球时,除了血细胞和大分子的朊外,血浆中的一部分水、电解质和有机物通过肾小球滤过膜进入肾小球囊腔内,形成滤过液,称之为原尿。
说明:原尿是血浆流经肾小球时通过肾小球的滤过作用而产生的,此然原尿是由血浆滤过而来,那么在具有肾血流量的前提下,还必须具有可通透的滤过膜和需要一定的压力。
原尿的生成必须具备两个条件:一是滤过膜的通透性;二是有效滤过压。
1.滤过膜的通透性
滤过膜:肾小球毛细血管官腔与肾小囊囊腔之间的膜,称滤过膜。组成:
肾小球毛细血管的内皮细胞、基膜和肾小囊的脏层细胞。三者紧贴在一起,形成有通透性的膜,总厚度<1um。
2.有效滤过压(滤过的动力)是存在于滤过膜两侧的压力差。滤过膜两侧均有促进和阻止滤过的力量:肾小球毛细血管一侧,血压促进滤过,血浆胶体渗透压阻止滤过;肾小球囊腔一侧,滤过液(原尿)胶体渗透压促进滤过,滤过液静压(肾球囊内压)阻止滤过。由于滤过液中蛋白含量很少,可忽略不计。
肾小球的有效滤过压=肾小球毛细血管血压-(血浆胶体渗透压+肾小球囊内压)
(二)肾小管和集合管的重吸收作用
原尿进入肾小管后称为小管液。小管液在流经肾小管和集合管的过程中,量和质都要发生很大变化:小管液中全部氨基酸、葡萄糖(包括漏出的少量蛋白质),大部分水、无机盐(钠、钾、氯、HCO3-等),部分其它物质(如氨、肌酐等),被重吸收入血液,剩余不到原尿1%的液体从肾乳头管流入肾盂,这就是终尿,也就是我们通常所说的尿液。几个概念:
- 重吸收:指物质从肾小管液中转运回血液的过程。
- 分泌:肾小管和集合管的上皮细胞将本身产生的物质或血液中的物质转运至管腔中的过程。
- 被动转运:指物质顺电化学梯度通过上皮细胞的过程。
- 主动转运:物质逆电化学梯度进行转运的过程。
各段肾小管和集合管的转运分述如下:
1.近球小管小管液中全部葡萄糖、氨基酸,85%的HCO3-,67%的Na+、Cl-、K+和水在近球小管中被重吸收,而且不受神经体液的调节。
钠泵是近球小管重吸收的关键动力。小管液中的钠和细胞内的H+与细胞膜上的交换体结合进行逆向转运,结果Na+顺浓度梯度进入细胞内,细胞内的H+分泌到小管腔中。Na+-H+的逆向转运现象又称Na+-H+交换,有利于HCO3—的重吸收。
小管液中的HCO3—不易透过管腔膜,HCO3—的重吸收是以CO2的形式变相吸收的。血液中的HCO3—是以钠盐NaHCO3的形式存在,在小管液中电离成Na+和HCO3—。小管上皮细胞内有碳酸酐酶,使CO2和H2O生成H2CO3,后者解离出H+和HCO3—。通过钠氢交换,小管液中的Na+进入细胞,与细胞内的HCO3—同向转运回血液(图7-5)。小管液中的HCO3—在碳酸酐酶作用下,与H+生成H2CO3,再分解产生水和CO2。CO2是脂溶性的,可以自由进入小管细胞。总之,钠泵不断将小管细胞内的Na+泵向周围组织,促进了钠氢交换,氢的分泌带动了HCO3—的重吸收(以CO2的形式)。有些药物(如乙酰唑胺)对碳酸酐酶有抑制作用,H+的生成减少,影响钠氢交换,NaHCO3重吸收随之减少。
在近球小管后半段,NaCl通过细胞旁路被重吸收。由于HCO3—是以CO2的形式重吸收的,而CO2是脂溶性的,易透过管腔膜,因此HCO3—的重吸收比Cl—快。在近球小管后半段Cl—的浓度比管周组织高20~40%。Cl—顺浓度梯度经细胞旁路(通过紧密连接进入细胞间隙)被动重吸收。Cl—带负电荷,Cl—重吸收产生的电位差促进Na+沿细胞旁路被动重吸收。
原尿中的K+约67%在近球小管重吸收,尿中的K+主要由远曲小管和集合管分泌。小管液中K+的浓度(4mmol/L)明显低于细胞内(150mmol/L),据此认为K+是逆浓度梯度重吸收的,但其机理尚不清楚。
肾糖阈近球小管对葡萄糖的重吸收有一定限度。当血中葡萄糖的浓度超过一定范围时(160~180mg/100ml)时,部分肾小管对葡萄糖的吸收达到极限而使尿中出现糖,此时的血糖浓度称为肾糖阈(renal glucose threshold)。
2.髓袢髓袢升支粗段对水不通透,所以NaCl的重吸收造成小管液低渗,组织液高渗。这种水和盐重吸收的分离有利于尿液的浓缩和稀释。速尿、利尿酸等利尿剂能作用于髓袢升支粗段,影响Na+:2Cl-:K+同向协同转运,干预尿液的浓缩而表现出利尿效应。
3.远曲小管和集合管远曲小管初段对水的通透性很低,但仍主动重吸收NaCl,继续产生低渗小管液。远曲小管后半段和集合管含两类细胞:主细胞和闰细胞。前者重吸收钠和水,后者分泌H+。在这里钠的重吸收仍然是靠钠泵逆电化学梯度主动进行的。
尿中钾的排泄量随摄入量的多少而相应变化。K+的分泌与钠的重吸收有密切关系。①钠主动重吸收是生电性的,使管腔内带负电位。这为K+的分泌提供了动力。②主细胞内K+浓度明显高于小管液,K+可顺浓度梯度经管腔膜上的K+通道扩散到小管液。
闰细胞分泌H+是一个逆电化学梯度的主动转运过程。有人认为管腔膜上有H+泵,能将细胞内的H+泵入小管腔中。分泌的H+与小管液中的HPO42—结合形成可滴定酸H2PO4—;与上皮细胞分泌的NH3结合形成NH4+。可滴定酸和NH4+均不易透过管腔膜而留在小管液中,成为尿液酸碱度的决定因素。
远曲小管和集合管在代谢过程中不断产生NH3(由谷氨酰胺脱氨而来),NH3是脂溶性的,能通过细胞膜向组织液和小管液中自由扩散。扩散速度取决于两侧液体的pH值。由于肾小管的泌氢作用,小管液中pH值较低,H+与分泌出来的NH3结合形成NH4+,降低了小管液中NH3的浓度,从而促进了NH3的分泌。
第三节尿液的浓缩和稀释
原尿可以看作是不含朊的血浆,其渗透压也与血浆相近,但排出的尿液有时渗透压比血浆高,称高渗尿,有时比血浆低,称低渗尿,有时是等渗的,称等渗尿。高渗尿说明尿被浓缩,而低渗尿说明尿被稀释,机体可根据不同的状况,排不同的尿,说明肾有浓缩和稀释的能力,尿是如何被浓缩和稀释的?
一、条件
1、肾髓质的组织间液经常处于高渗状态,并具有渗透压梯度,即从肾皮质到髓质,渗透压逐渐增加,越接近乳头部渗透压越高。
- 原尿由近曲小管到髓袢到远曲小管再到集合管的过程中,各部位对原尿中溶质和水的通透性不同。
二、尿液的浓缩和稀释过程
渗透压梯度与尿液浓缩有密切关系,集合管与髓袢平行,处于渗透压梯度中,当低渗尿从皮质集合管向乳头管方向流动时,抗利尿激素分泌较多,使集合管上皮细胞对水的通透性加大,水因管外的渗透压梯度而逐渐被吸出,于是被浓缩成高渗尿。
当机体内水分过多时,抗利尿激素分泌减少,使集合管和远曲小管壁不易透过水,从髓袢升支进入远曲小管的低渗液由于Na+在远曲小管和集合管中被主动重吸收而水确难于吸收,其渗透压进一步降低,因而形成低渗尿。
如前所述,髓袢升支粗段能主动重吸收Na+、Cl—而对水不通透,使小管液在此变为低渗液。此低渗液流经远曲小管和集合管时,Na+、Cl—继续被重吸收,而水的通透性则受抗利尿激素 的调节。体内水过剩时,抗利尿激素 分泌减少,远曲小管和集合管对水的通透性很低,水的重吸收很少,结果就排出低渗尿。机体缺水时,抗利尿激素 分泌增多,远曲小管和集合管对水的通透性增高,由于管周髓质的渗透压很高,在渗透压差作用下,水被大量重吸收,从而形成高渗尿。
第四节尿生成的调节
尿的生成有赖肾小球的滤过和肾小管、集合管的重吸收与分泌作用。尿生成的调节也发生在这些环节。
一、影响肾小球滤过的因素
1.滤过膜的通透性及滤过面积正常情况下,滤过膜的通透性和滤过面积相对稳定,对滤过影响不大。病理情况下,如急性肾小球肾炎时,肾小球毛细血管管腔变窄或阻塞,有滤过功能的肾小球减少,滤过膜增厚通透性降低,滤过面积减少,滤过率降低,出现少尿或无尿。
2.肾小球毛细血管血压肾血流有自身调节作用,当动脉血压在正常范围波动时,肾小球毛细血管血压维持稳定,滤过率基本不变。大失血等,血压明显下降,肾小球毛细血管血压也相应下降,有效滤过压降低。高血压晚期,入球小动脉硬化而缩小,肾小球毛细血管血压可明显降低,有效滤过压减小导致少尿。
3.肾小球囊内压正常情况下囊内压较稳定。肾结石、肿瘤或其它原因引起输尿管阻塞;某些药物在肾小管中析出;溶血过多,血红蛋白堵塞肾小管等,都会导致囊内压升高而使滤过率降低。
4.血浆胶体渗透压血浆胶体渗透压一般变化不大。静脉快速注入生理盐水时,可使血浆胶体渗透压减小,滤过率增大。
5.肾血浆流量这主要影响滤过平衡的位置。肾血浆流量增大,肾小球毛细血管内血浆胶体渗透压的上升速度就慢,滤过平衡就靠近出球小动脉,有效滤过压和滤过面积就增加,滤过率随之增加;反之则减小。严重缺氧、中毒性休克时,交感神经兴奋,肾血流量和血浆流量显着减少,滤过率也减小。
二、影响肾小管和集合管重吸收及分泌作用的因素
(一)小管液中溶质的浓度
水由低渗溶液向高渗溶液扩散。肾小管内溶质浓度越高,其渗透压也越高,越不利于水的重吸收,结果尿量就会增加。糖尿病患者,渗透性利尿剂(如甘露醇)都是因小管液浓度增加而导致尿液增多的。
(二)球—管平衡
近球小管对溶质和水的重吸收量不是固定不变的,而随肾小球滤过率的变动而变化。滤过率增大,滤液中水、钠增多,重吸收随之增加;反之亦然。实验表明,无论肾小球滤过率增加还是减少,近球小管是定比重吸收的,即近球小管的重吸收率始终占肾小球滤过率的65~70%,这种现象称球—管平衡(glomerulotubular balance)。其意义在于使尿液的质和量不因肾小球滤过率的增减而出现大幅度的变动。
定比重吸收的机制与管周毛细血管血压和胶体渗透压的变化有关。在肾血流量不变的前提下,肾小球滤过率增加,进入近球小管旁毛细血管的血量就减少,血压下降,而胶体渗透压升高。这会促进小管旁组织间液进入毛细血管,使组织间隙静水压下降,通过紧密连接返回小管腔的回漏量减少,从而使小管液的重吸收量增加,仍可达滤过率的65~70%。当滤过率减少时发生相反的变化,使重吸收率保持相对稳定。
球—管平衡在某些情况下会遭破坏,如根皮苷中毒时近球小管对葡萄糖的重吸收能力减弱,出现渗透性利尿。
(三)管—球反馈
肾血流量和肾小球滤过率增加或减少时,到达远曲小管致密斑的小管液的流量随之增减,致密斑能感受小管液中NaCl含量的改变,发出信息,使肾血流量和肾小球滤过率恢复正常。小管液流量变化影响肾血流量和滤过率的现象称管—球反馈。
(四)抗利尿激素
抗利尿激素是由下丘脑的视上核和室旁核的神经细胞分泌的9肽激素,经下丘脑—垂体束到达神经垂体后释放出来。其主要作用是提高远曲小管和集合管对水的通透性,促进水的吸收,是尿液浓缩和稀释的关键性调节激素。此外,该激素还能增强内髓部集合管对尿素的通透性。
调节抗利尿激素分泌的主要因素有晶体渗透压、循环血量和动脉血压。机体因某种原因失水时,晶体渗透压升高,引起抗利尿激素 分泌增多,使尿量减少尿液浓缩。相反,大量饮清水后,血液稀释,晶体渗透压降低,抗利尿激素 分泌减少,肾对水的重吸收减少,结果排出大量低渗尿,将体内多余的水排出体外,此现象称水利尿(water diuresis)。循环血量增加,左心房扩展,刺激容量感受器,通过迷走神经将信息传到中枢,抑制下丘脑—垂体后叶释放抗利尿激素 ,尿量增加。血量减少时,出现相反的变化。动脉血压升高时,刺激颈动脉窦压力感受器,反射性地抑制抗利尿激素 的释放。
(五)肾素—血管紧张素—醛固酮系统
肾素是由近球小体中的颗粒细胞分泌的一种蛋白水解酶,可将血浆中的血管紧张素原转变为血管紧张素I,后者经血液循环(特别是经过肺),降解成血管紧张素II,从而刺激肾上腺皮质球状带合成和分泌醛固酮。
血管紧张素II对尿生成的调节:①刺激醛固酮的合成和分泌,调节K+、Na+转运。②直接刺激近球小管对NaCl的重吸收,减少其排泄。③刺激抗利尿激素 的释放,使尿量减少。
醛固酮对尿生成的调节:促进远曲小管和集合管的主细胞重吸收Na+而排出K+,有保钠排钾作用。醛固酮的分泌除受血管紧张素调节外,血K+浓度升高和血Na+浓度降低,都能直接刺激肾上腺皮质分泌醛固酮。
(六)甲状旁腺激素和降钙素
甲状旁腺分泌的甲状旁腺激素对肾脏有如下作用:抑制近球小管对磷酸盐的重吸收,促进其排泄;促进远曲小管和集合管对Ca2+的重吸收,使尿钙减少;增强肾小管细胞内羟化酶的活性,使25-(OH)D3转变为1,25-(OH)2D3。
甲状旁腺C细胞分泌的降钙素对肾脏的作用:抑制肾小管对钙、磷的重吸收,促进钙、磷排泄;抑制肾脏羟化酶的活性,阻止25-(OH)D3的活化。
(七)心房利尿钠肽
心房利尿钠肽是由心房肌分泌的28肽激素,有明显促进NaCl和水排出的作用。
此外,交感神经兴奋时通过引起入球、出球小动脉收缩,刺激近球小体颗粒细胞分泌肾素,增加近球小管和髓袢对Na+、Cl—和水的重吸收,使尿量减少。
肾脏除完成泌尿功能,分泌肾素,活化25-(OH)D3外,还能分泌促红细胞生成素和前列腺素。由肾系膜细胞分泌的促红细胞生成素主要作用于骨髓,刺激红细胞的生成。肾髓质乳头部能合成前列腺素PGA2和PGE2,引起肾(皮质)局部血管扩张,肾血流增加。
第五节排尿
尿由肾脏生成后,经肾盂到输尿管,借输尿管的蠕动,流入膀胱,贮积到相当容量时排出体外的过程称排尿。是反射动作。
一、膀胱与尿道的神经支配
1、盆神经-副交感神经,兴奋→ 膀胱逼尿肌收缩,膀胱内括约肌松弛 → 促进排尿
2、腹下神经-交感神经 。传出F兴奋时,使膀胱逼尿肌松弛,膀胱内括约肌收缩→ 抑制排尿(贮尿)
3、阴部神经-躯体神经。直接受意识和反射的控制,兴奋时,可使膀胱外括约肌收缩,抑制排尿。
二、排尿反射
当膀胱内的尿液充盈到一定程度时(人400-500ml),膀胱内压升高,刺激了膀胱壁的牵张感受器,冲动沿盆神经感觉纤维传入腰荐部脊髓,再沿脊髓上传到脑干和大脑皮层的高级中枢,产生尿意。如果条件允许,兴奋→排尿;不允许 → 抑制 → 不排尿。排尿结束后,引起排尿反射的刺激因素被解除,初级中枢在高级中枢的调控下受到抑制,逼尿肌紧张性减弱,内外括约肌紧张性加强,膀胱又进入蓄尿状态。
尿的生成和排尿是两个既相对独立又相互影响、协调统一的过程,任何一方发生异常都会对泌尿活动造成不同程度的影响。
第九章 内分泌
第一节概述
一、内分泌器官和内分泌细胞
内分泌系统:是由内分泌腺和分散存在于某些器官组织中的内分泌细胞组成的一个体内信息传递系统。
内分泌(endocrine):内分泌腺或内分泌细胞合成和分泌的特殊化学物质,通过血液循环或扩散传递给相应的靶细胞,调节其生理功能的过程,称为内分泌(endocrine)。
内分泌器官:具有典型的腺体结构,且没有导管,其分泌物直接进入血液或组织液,所以又被称为内分泌腺。内分泌腺包括垂体、甲状腺、甲状旁腺、肾上腺、胰岛、性腺、松果腺和胸腺等。
二、激 素(hormone)
(一)激素的概念
由动物的内分泌腺或散在的内分泌细胞所分泌的高效能的生物活性物质,经组织液或血液传递而发挥其调节作用,这种化学物质称为激素。
(二)激素的传递方式
根据激素到达靶细胞的途径,其传递有如下几种方式
1.远距分泌(telecrine)大多数激素进入血液,经血液循环到达远距离的靶细胞而发挥生理作用,即传统的内分泌概念。
2.旁分泌(paracrine)某些激素进入细胞间液,通过扩散到达邻近的靶细胞起作用。
3.自身分泌(antocrine)内分泌细胞分泌的激素在局部扩散,又返回作用于该内分泌细胞而发挥反馈作用。
4.神经分泌(neurocrine)神经细胞分泌的化学信使(神经递质)由轴突末梢释放,经突触传递至突触后细胞影响其生理功能。
5.神经内分泌(neuroendocrine)神经细胞分泌的化学信使(激素)进入血液,通过血流到达靶细胞起作用。
此外,激素还可通过细胞间的微管连接到达另一细胞起调节作用,称为表分泌(epicrine)。
(三)激素的分类
激素的种类繁多,来源复杂,按其化学性质可分为含氮激素(nitrogenous hormone)和类固醇(甾体)激素(steroid hormone)两大类。
1.含氮激素
(1)肽类和蛋白质激素主要有下丘脑调节肽、腺垂体激素、神经垂体激素、胰岛激素、甲状旁腺激素、降钙素以及胃肠激素等。
(2)胺类激素包括肾上腺素、去甲肾上腺素和甲状腺激素等。
2.类固醇(甾体)激素类固醇激素是由肾上腺皮质和性腺分泌的激素。另外,胆固醇的衍生物—1,25-二羟胆钙化醇(又称1,25-二羟维生素D3)也被作为激素看待。
(四)激素的一般特性
1.激素的信息传递作用内分泌系统以化学形式传递信息,即依靠激素在细胞与细胞之间进行信息传递。
激素作为体内的“信使”,其作用主要包括:①调节物质代谢,维持代谢的稳态。②调节细胞的分裂和分化,保证机体的正常生长、发育和成熟,并影响衰老过程。③影响中枢神经系统和植物性神经系统的发育和功能。④调节生殖器官的发育和成熟,并保证生殖过程正常进行。⑤与神经系统密切配合,使机体更有效地适应内外环境的变化。
2.激素作用的相对特异性进入体液中的激素只选择地作用于某些器官、组织和细胞,这称为激素作用的特异性。被激素选择作用的器官、组织和细胞,分别称为靶器官(target organ)、靶组织(target tissue)和靶细胞(target cell)。
3.激素的高效能生物放大作用激素在血液中的浓度都很低,一般在纳摩尔(nmol/L),甚至在皮摩尔(p mol/L)数量级,但其作用显著。这是由于激素与靶细胞的受体结合后,细胞内发生一系列酶促放大作用,一个接一个,逐级放大效果,形成一个效能极高的生物放大系统。
4.激素间的相互作用当多种激素共同参与某一生理活动的调节时,激素与激素之间往往存在着协同作用或颉颃作用,这对维持其功能活动的相对稳定起着重要作用。
另外,有的激素本身并不能直接对某些器官、组织或细胞产生生理效应,然而在它存在的条件下,可使某一种激素的作用明显增强,即对另一种激素的调节起支持作用,这种现象称为允许作用(permissive action)。
(五)激素的合成、分泌、转运和代谢
1.激素的合成先在细胞核内将DNA中的遗传信息转录到mRNA,然后在胞浆中通过mRNA的翻译过程形成多肽激素链。
2.激素的分泌内分泌细胞合成激素后释放到细胞外或血液中的过程,称为分泌(secretion)。
3.激素的转运激素分泌后经血液循环或体液扩散到靶细胞的过程,称为激素的转运。
4.激素的代谢激素从释放出来直到失活并被消除的过程,称为激素的代谢。代谢的速度通常以半衰期(half-life period),
半衰期:即激素的浓度或活性在血液中减少一半所需要的时间。
三、激素作用的机制
(一)含氮激素的作用机制—第二信使学说(second messenger hypothesis)
这个学说的主要内容包括:
①激素作为第一信使,首先与靶细胞膜上具有立体构型的专一性受体结合;
②激素与受体结合后,激活膜上的腺苷酸环化酶(AC)系统;
③ 在Mg2+存在的条件下,AC促使ATP转变为环一磷酸腺苷(cAMP),cAMP是第二信使,信息由第一信使传递给第二信使;④cAMP使无活性的蛋白激酶A (PKA)激活。PKA具有两个亚单位,即调节亚单位(PKr)与催化亚单位(PKc)。cAMP与PKc结合,导致PKr与PKc脱离而使PKA激活,催化细胞内多种蛋白质发生磷酸化反应,包括一些酶蛋白发生磷酸化,从而引起靶细胞各种生理生化反应(图10-2)。cAMP发挥作用后,迅速被细胞内的磷酸二酯酶(PDE)降解为5′-磷酸腺苷(5′-AMP),而失去活性。
近年来的研究资料表明,cAMP并不是唯一的第二信使,可能作为第二信使的化学物质还有环一磷酸鸟苷(cGMP)、三磷酸肌醇(IP3)、二酰甘油(DG)、Ca2+等。
现概述如下:
1.激素与受体的相互作用
由于激素和受体可以相互诱导而改变本身的构型以适应对方的构型,所以激素与受体能够发生专一结合。激素与受体之间的结合力称为亲和力(affinity)。
2.G蛋白在信息传递中的作用
激素受体与腺苷酸环化酶是细胞膜上两类分开的蛋白质。激素受体结合的部分在细胞膜的外表面,而腺苷酸环化酶在膜的胞浆面,在两者之间存在一种起耦联作用的调节蛋白—鸟苷酸结合蛋白,简称G蛋白。G蛋白由α、β和γ三个亚单位组成,α亚单位上有鸟苷酸结合位点。当G蛋白上结合的鸟苷酸为GTP时则激活而发挥作用,但当G蛋白上的GTP水解为GDP时则失去活性。当激素与受体结合时,活化的受体便与G蛋白的α亚单位结合,并促使其与β、γ亚单位脱离,才能对腺苷酸环化酶起激活或抑制作用。
3.三磷酸肌醇和二酰甘油为第二信使的信息传递系统
这一学说认为,在激素的作用下,可能通过G蛋白的介导,激活细胞膜内的磷脂酶C(PLC),它使由磷脂酰肌醇二次磷酸化生成的磷脂酰二磷肌醇(PIP2)分解,生成IP3和DG。DG生成后仍留在膜中,IP3则进入胞浆。
IP3的作用是促使线粒体和内质网中的Ca2+释放进入胞浆,随后Ca2+与细胞内的钙调蛋白(calmodulin, CaM)结合,激活蛋白激酶,促进蛋白质磷酸化,从而调节细胞的功能活动。
DG的作用主要是它能特异性激活蛋白激酶C(PKC),PKC的激活依赖于Ca2+的存在。激活的PKC与PKA一样可使多种蛋白质或酶发生磷酸化反应,进而调节细胞的生物效应。另外,DG的降解产物花生四烯酸是合成前列腺素的原料,花生四烯酸与前列腺素的过氧化物又参与鸟苷酸环化酶的激活,促进cGMP的生成。cGMP作为另一种可能的第二信使,通过激活蛋白激酶G(PKG)而改变细胞的功能。
(二)类固醇激素作用机制—基因表达学说
该学说认为:类固醇激素的分子小(分子量仅为300左右)、呈脂溶性,因此可透过细胞膜进入细胞。在进入细胞之后,经过两个步骤影响基因表达而发挥作用,故把此种作用机制称为二步作用原理,或称为基因表达学说。
1、第一步是激素与胞浆受体结合,形成激素—胞浆受体复合物。当激素进入细胞内与胞浆受体结合后,受体蛋白发生构型变化,从而使激素—胞浆受体复合物获得进入核内的能力,由胞浆转移至核内。
2、第二步是与核内受体结合,形成激素—核受体复合物,从而激发DNA的转录过程,生成新的mRNA,诱导蛋白质合成,引起相应的生物效应(图10-3)。
(三)激素作用的其它方式
也可以进入细胞核与核受体结合,然后通过基因表达,促进或抑制细胞的代谢;
类固醇激素还可直接作用于细胞膜的类脂成分,引起细胞膜的通透性变化等。
四、激素分泌的调控
(一)神经调节
内外环境发生变化的信号传入中枢神经系统经整合后可直接或间接地影响和调节激素的分泌。
例如神经垂体、肾上腺髓质和某些胃肠道内分泌细胞激素的分泌,直接受神经控制;吮吸动作或扩张阴道可反射性引起神经垂体释放催产素;动物在紧急情况下引起儿茶酚胺类激素大量释放,等等。
(二)体液调节
1.激素的反馈调节
当一种激素分泌后,经体液作用于靶细胞引起特异的生理效应时,几乎同时反馈控制该激素的分泌。引起激素分泌减少的称负反馈,引起分泌增加的称为正反馈,根据反馈路线长短,有长反馈(long-loop feedback)、短反馈(short-loop feedback)和超短反馈(ultrashort-loop feedback)三种。
2.代谢物的反馈调节例如胰岛β细胞分泌胰岛素调节血糖浓度,而血糖浓度升高或降低则可促进或抑制胰岛素的分泌。
第二节下丘脑的内分泌功能
一、下丘脑的神经内分泌细胞
下丘脑的神经内分泌细胞能分泌神经肽或肽类激素,所以又称为肽能神经元(peptidergic neuron)。
下丘脑的神经内分泌细胞可分为大细胞和小细胞两种。
(1)大细胞主要分布在下丘脑视前区的视上核和室旁核。
(2)小细胞分散存在于下丘脑底部视前区的室旁核、视上核、结节区的弓状核、腹内侧核内,它们的轴突主要组成结节-垂体束,终止于正中隆起和漏斗柄。这些细胞释放的激素都作用于腺垂体,引起(或抑制)腺垂体激素的分泌,故将这些细胞所在的区域称为下丘脑促垂体区(hypothalamic hypophysiotropic area)。
二、下丘脑与垂体的功能联系
下丘脑与垂体之间的功能联系有两种方式:
一是通过垂体门脉,在正中隆起或垂体柄部与垂体门脉的初级毛细血管网相接,神经分泌物从这里释放进入血液,再沿门脉血管到达腺垂体,形成次级毛细血管网,与腺垂体细胞相接。
另一种联系方式是直接的神经联系,即通过下丘脑-垂体束,下丘脑的神经分泌物沿轴突的轴浆流动运送到神经垂体。所以,下丘脑与垂体一起组成下丘脑-垂体功能单位。
三、下丘脑分泌的激素(因子)及其生理作用
(一)促甲状腺激素释放激素
促甲状腺激素释放激素(TRH)是由谷氨酸、组氨酸和脯氨酸结合而成的3肽。
TRH的主要生理作用:促进腺垂体的促甲状腺激素细胞合成和分泌促甲状腺激素,从而促进甲状腺激素的合成和释放。还轻度刺激腺垂体分泌催乳素和生长激素。
大脑和脊髓,也有TRH存在,其作用可能与神经信息传递有关。
(二)促性腺激素释放激素
促性腺激素释放激素(GnRH)是10肽激素。
主要生理作用:促进腺垂体的促性腺细胞合成和释放卵泡刺激素和黄体生成素两种促性腺激素,但对黄体生成素的刺激作用较强,所以又称黄体生成素释放激素(LRH)。
(三)生长激素释放激素
生长激素释放激素(GHRH)是多肽激素。
生理作用:促进腺垂体生长激素细胞合成和分泌生长激素。
(四)生长抑素
生长抑素(生长激素释放抑制激素,GIH或somatostatin)是由116个氨基酸的大分子肽裂解而来的14肽。
主要作用:抑制腺垂体生长激素的基础分泌,也抑制腺垂体对多种刺激所引起的生长激素分泌反应,包括运动、进食、应激、低血糖等。另外,生长抑素还可抑制黄体生成素、卵泡刺激素、促甲状腺激素、催乳素及促肾上腺皮质激素的分泌。
(五)促肾上腺皮质激素释放激素
促肾上腺皮质激素释放激素(CRH)为41肽。
主要作用:促进腺垂体合成与释放促肾上腺皮质激素。
(六)其它的下丘脑调节肽
催乳素释放因子(PRF)促使催乳素分泌。
乳素释放抑制因子(PIF)抑制催乳素分泌。
促黑素细胞激素释放因子(MRF)。
促黑素细胞激素释放抑制因子(MIF)可能是催产素裂解出来的两种小分子肽。
MRF促进促黑激素的释放,而MIF则抑制促黑激素的释放。
四、下丘脑调节肽分泌的调节
下丘脑调节肽的分泌,受两方面的调控:
一方面,下丘脑促垂体区细胞的分泌作用受到脑内其它部位(主要是中脑、边缘系统和大脑皮质)传来的神经纤维所释放的神经递质如脑啡肽、内啡肽和P物质等肽类物质,以及多巴胺、5-羟色胺和去甲肾上腺素等单胺物质的调节作用。
另一方面,下丘脑调节肽的分泌受体液中激素和代谢物浓度的反馈调节作用,特别是靶腺激素的反馈调控。
第三节垂体
一、腺 垂 体
腺垂体是体内最重要的内分泌腺。它由不同的腺细胞分泌七种激素:
1、生长激素:由生长激素细胞分泌;
2、促甲状腺激素:由促甲状腺激素细胞分泌;
3、促肾上腺皮质激素(adrenocorticotropic hormone, ACTH)与促黑(素细胞)激素( melanophore-stimulating hormone, MSH):由促肾上腺皮质激素细胞分泌;
4、卵泡刺激素(follicle-stimulating hormone, FSH)与黄体生成素(luteinizing hormone,LH):由促性腺激素细胞分泌;
5、催乳素(prolactin,PRL):由催乳素细胞分泌。在腺垂体分泌的激素中,TSH、ACTH、FSH与LH均有各自的靶腺,分别形成:①下丘脑-垂体-甲状腺轴。②下丘脑-垂体-肾上腺皮质轴。③下丘脑-垂体-性腺轴。腺垂体的这些激素是通过调节靶腺的活动而发挥作用的。而GH、PRL与MSH则不通过靶腺,分别直接调节个体生长、乳腺发育与泌乳、黑素细胞活动等。所以,腺垂体激素的作用极为广泛而复杂。
(一)腺垂体激素的生物学作用
1.生长激素(growth hormone,GH) GH是由一条肽链构成的单纯蛋白质。各种动物的GH大约由190个氨基酸组成,分子量约为20000~26000,GH的生理作用是促进物质代谢与生长发育,对机体各个器官与各种组织均有影响,尤其对骨骼、肌肉及内脏器官的作用最为显著。
(1)促进生长作用由于GH能促进骨、软骨、肌肉以及其它组织分裂增殖、蛋白质合成增加,从而增加细胞的体积和数量。生长介素主要的作用是促进软骨生长,
(2)促进代谢作用GH可通过生长介素促进氨基酸进入细胞,加速蛋白质合成,包括软骨、骨、肌肉、肝、肾、心、肺、肠、脑及皮肤等组织的蛋白质合成增强;GH促进脂肪分解,增强脂肪酸氧化;提高血糖水平。
2.促甲状腺激素(thryoid-stimulating hormone,TSH)TSH是一种糖蛋白,分子量约为25000。它能促进甲状腺细胞增生及其活动,促使甲状腺激素的合成和释放。
3.促肾上腺皮质激素(adrenocorticotropic hormone,ACTH)ACTH是含有39个氨基酸的多肽,分子量约为4500。主要作用是促进肾上腺皮质的发育以及糖皮质激素的合成和释放。在应激时,ACTH分泌增加。
4.促性腺激素(gonadotropic hormone, ganadotropin, GTH)GTH是一种糖蛋白,分子量约为30000。GTH可分为卵泡刺激素(FSH)和黄体生长素(LH)两种。
(1)FSH在LH的协同作用下,FSH对卵巢的作用是促进卵泡细胞增殖和卵泡生长,并引起卵泡液分泌;对睾丸的作用是作用于曲精细管的生殖上皮,促进精子的生成,并在睾酮的协同作用下使精子成熟。
(2)LH在FSH的协同作用下,LH对卵泡有明显的促生长作用,并促进卵泡雌激素合成和卵泡成熟,并激发排卵。排卵后的卵泡在LH作用下转变成黄体。
5.催乳素(prolactin,PRL)PRL是一种单纯蛋白质激素。
6.促黑素细胞激素(MSH)MSH的主要作用是刺激黑素细胞内黑素的生成和扩散,使皮肤颜色变暗、变黑。
(二)腺垂体激素分泌的调节
腺垂体的分泌功能一方面受中枢神经系统,特别是下丘脑的控制,另一方面也受外周靶腺所分泌的激素和代谢产物的反馈调节,在下丘脑-腺垂体—靶腺轴多激素间的相互作用下,使靶腺激素在血液中保持动态平衡。
下丘脑的调控表现在以下三个面:
①下丘脑促垂体区释放激素和释放抑制激素的作用:GTH、TSH、ACTH的分泌直接受下丘脑分泌的相应的释放激素的控制;而GH、PRL和MSH则分别受下丘脑释放的释放激素和释放抑制激(因子)的双重控制。
②神经肽、神经递质和神经调制物的作用:升压素、神经降压肽、P物质、阿片样肽、5-羟色胺等可促进GH分泌;肾上腺素、去甲肾上腺素、γ-氨基丁酸、5-羟色胺等对MSH和ACTH分泌有调节作用。
③其他中枢部位和外周感受器的作用:MSH分泌还受下丘脑的直接控制;吮吸刺激乳头可反射地引起PRL分泌增加。
2.反馈调节激素在血液中的浓度通过反馈途径可直接影响,也可间接通过下丘脑影响腺垂体激素的分泌。血液中甲状腺激素和皮质醇浓度的下降既可直接作用于腺垂体,也可通过对下丘脑释放激素的改变间接作用于腺垂体,从而使TSH和ACTH分泌加强。相反,甲状腺激素或皮质醇升高可通过同样途径引起TSH或ACTH分泌减少(图10-6)。
二、神经垂体
神经垂体主要由下丘脑-垂体束的无髓神经末梢与神经胶质细胞分化而成的神经垂体细胞所组成,是贮存与释放下丘脑视上核和室旁核的神经分泌物的部位。因此,可以把神经垂体看作是下丘脑的延伸部分。
(一)神经垂体激素的化学
神经垂体释放两类激素,一类是升压素(vasopressin,VP),又称抗利尿激素(antidiuretic hormone,ADH);另一类是催产素(oxytocin,OXT)。升压素和催产素都是9个氨基酸组成的9肽,结构大致相同,分子量约1000。由于分子结构中N-端第1和第6位的半胱氨酸以二硫键相连接成一分子胱氨酸,所以常把这两类激素看作是8肽。哺乳动物的升压素有两种类型:8-精升压素和8-赖升压素。这些激素的化学结构如图10-7:
(二)神经垂体激素的生理作用
1.催产素催产素的主要作用是促进子宫肌收缩,促使胎儿、胎衣排出及产后止血。
2.升压素升压素的主要生理作用是调节血浆渗透压、血容量和血压。由于其促使肾远曲小管和集合管对水的重吸收,因而减少尿量,具有抗利尿效应,因此又称为抗利尿激素。
(三)神经垂体激素分泌的调节
1.升压素分泌的调节在下丘脑有渗透压感受器,当血浆渗透压增加1~2%,渗透压感受器兴奋,冲动沿下丘脑-垂体束传至神经垂体,引起升压素释放,发挥保水作用,以维持渗透压平衡。与此同时,在下丘脑视前区附近有另一渗透压感受器称为饮水中枢,饮水中枢与视上核的渗透压感受器共同作用,调节因脱水带来的渗透压变化。当血浆晶体渗透压下降,则发生逆反应。
2.催产素分泌的调节催产素的释放主要依靠来自子宫颈,阴道和乳房的一些刺激,当交配、分娩或吮乳时,都可反射性地引起催产素的释放。与此相关的各种条件刺激,如幼仔出现、喊叫、挤乳准备还可引起催产素的条件反射性释放。
第四节甲 状 腺
位置:颈部气管的两侧。人位于喉头的前下方,分左右两叶;动物:喉和气管,1、2气管环的两侧,中间有一峡部相连。
一、甲状腺激素
特点:分泌的激素含碘,甲状腺含碘总量伟5000ug,甲状腺能聚集碘量90%以上,可合成甲状腺素T4、T3。
1、甲状腺激素的贮存、释放、运输与代谢
结构:是酪AA的碘化产物
合成:(1)碘的聚积:甲状腺能逆浓度梯度把I-聚积到腺体内,这一机制称为“碘泵”(通过ATP对Na+、K+的转运主动运碘)。腺泡内I-浓度比血中高25-50倍。
(2)碘的氧化和酪AA碘化:
聚积的I-过氧化物酶I2 + 2e
酪AA残基上的氢原子被碘取代或碘化,首先出现一碘酪AA残基和二碘酪AA残基。
(3)耦联反应(缩合反应):两个二碘酪AA缩合成T4,一个一碘酪AA缩合成T3
贮存:合成后的T4、T3仍结合在甲状腺球蛋白上,贮存在腺泡腔中,贮存量很大,一般可供数周(1、2、3月)使用。
释放:甲状腺受到适宜刺激后或在TSH作用下经微绒毛的吞噬(胞饮)作用进入腺C,再被分解成T4、T3进入血液,分泌入血液中的T4:T3的比例一般为20-30:1,各种动物都以T4为主,而在血液中都有两种形式存在:游离型、结合型。
2、甲状腺激素的生理作用
- 生长发育
- 对NS的影响
- 对生殖器官的发育成熟和功能
- 对代谢的影响
- 对生长的影响
3、甲状腺激素分泌的调节
- 下丘脑-腺垂体-甲状腺轴的作用
- 自身调节
在完全缺乏TSH或者TSH浓度基本不变的情况下发生的一种调节称自身调节。甲状腺腺泡内有机碘的含量决定着甲状腺转运碘的能力,当有机碘的含量升高,腺体摄取和浓缩碘的能力降低,反之,当腺细胞内缺乏有机碘时,摄取和浓缩碘的能力增强,这样以适应食物中碘供应量的增减。
地方性甲状腺肿,食物少碘,T3、T4少,负反馈减弱,TRH与TSH分泌多,使甲状腺增生。
第五节甲状旁腺、维生素D3和甲状腺C细胞
甲状旁腺和甲状腺C细胞(滤泡旁细胞)分泌的激素以及1,25-二羟维生素D3[1,25-二羟胆钙化醇,1,25-(OH)2D3]在调节机体钙、磷代谢,维持血钙、血磷的动态平衡和保证神经、肌肉的正常兴奋性方面,起着重要作用。
一、甲状旁腺素(PTH)
一般有两对,呈豆粒状。
由两类细胞组成:主C,分泌PTT,84AA组成的单链多肽。
嗜酸性细胞,功能不清。
(一)甲状旁腺素的生理作用
甲状旁腺素的主要作用为维持血Ca2+恒定。
摘除甲状旁腺后,血Ca减少,血P增加,将导致动物死亡。
PTH对肾脏的作用是:
①促进肾小管对钙的重吸收,从而减少尿钙的排出。抑制肾小管对磷的吸收,从而增加尿磷的排出。
②促进肾小管内羟化酶的活性,使25-(OH)D3转变成1,25-(OH)2D3。使肾小管上皮内的Ca2+输送进入细胞外液,再进入血液。
(三)甲状旁腺素分泌的调节
PTH的分泌主要受调节物浓度的影响。
- 主要受血浆钙浓度的调节:血Ca维持在正常情况下,PTH也持续少量分泌。血Ca减少,PTH的分泌增加;血Ca增加,PTH的分泌减少,这也是一个负反馈过程。
- 1,25-(OH)2D3增加,PTH分泌减少。
- 降钙素大量释放时,可促进PTH的分泌作用,使血P增加,PTH分泌增加。
二、维生素D3
(一)维生素D3的活化
维生素D3(VD3)又称胆钙化醇(cholecalciferol),主要由皮肤中的7-脱氢胆固醇经日光中的紫外线照射转化而来。维生素D3并非是其生物活性形式,它需在肝脏先羟化成25-羟维生素D3[25-(OH)-D3],然后在肾脏进一步羟化转变成1,25-二羟维生素D3[1,25-二羟胆钙化醇,1,25-(OH)2D3],才有活性。
在肾脏形成的1,25-(OH)2D3经血液抵达肠道和骨,促进使钙磷吸收和骨钙动员,升高血钙水平。
(二)1,25-(OH)2D3的生理作用
1,25-(OH)2D3的主要生理作用是促进小肠上皮细胞对Ca2+的吸收,实现这一效应可能通过以下三个途径:①肠上皮细胞Ca2+的吸收速率与钙结合蛋白的数目成正比,1,25-(OH)2D3能促进钙结合蛋白质的合成。②促进上皮细胞刷状缘上钙激活的ATP酶的生成。③促进上皮细胞膜内碱性磷酸酶的形成,并维持酶的活性。
1,25-(OH)2D3对钙动员和骨盐沉积均有重要作用,投以大剂量维生素D3可动员骨钙入血,血钙浓度升高。在缺乏VD3的情况下,PTH对骨的作用明显减弱甚至消失。
(三)1,25-(OH)2D3生成的调节
25-(OH)D3在肾脏中转变1,25-(OH)2D3的过程主要受PTH的调节,血钙浓度低于正常水平时,PTH分泌增多,肾脏生成1,25-(OH)2D3的过程相应增强;血钙浓度升高后,PTH分泌减少,1,25-(OH)2D3生成也减少。
三、甲状腺C细胞
甲状腺C细胞可分泌降钙素(calcitonin, CT),降钙素是由32个氨基酸组成的多肽,分子量约3000,由于能降低血中Ca2+浓度而得名。
(一)降钙素的生理作用
主要是降低血钙和血磷,对幼畜尤其明显,它的靶器官是骨,对肾脏和肠道也有一定作用。
1.对骨的作用降钙素能抑制骨原始细胞转变成破骨细胞,又加速破骨细胞转化为成骨细胞。因此破骨细胞作用被抑制,溶骨过程减弱而成骨过程增强,使骨组织释放入血的钙盐减少,血钙和血磷浓度降低。
2.对肾脏的作用降钙素能抑制肾小管对钙、磷、钠、钾、铁和氯等离子的重吸收,使其在尿中的排出量增加。
3.对胃肠道的作用降钙素对胃肠道吸收钙没有直接作用,但能抑制肾内的25-(OH)-D3转变为1,25-(OH)2D3的过程,因而间接地的抑制肠道对钙的吸收。
(二)降钙素分泌的调节
1、降钙素的分泌受血钙浓度调节,血钙浓度升高时,降钙素分泌增多;反之则分泌减少。降钙素与PTH对血钙的作用相反,它们共同调节血浆钙离子浓度的相对恒定。两者的主要差别为:①降钙素分泌启动较快,而PTH分泌较慢;② 降钙素只对血钙水平产生短期调节作用,其作用很快被强有力的PTH作用所取代,后者对血钙浓度发挥长期调节作用。
2、近年发现,胃肠激素等可刺激CT的分泌,这样可防止食后血钙过高,可见血钙的维持要受PTH和CT的共同调节,它们之间是相互拮抗的。
第六节肾 上 腺
动物的肾上腺位于肾脏的前缘,人位于肾脏的上方,两个左右各一。
肾上腺髓质(内)儿茶酚胺激素
肾上腺
肾上腺皮质(外)类固醇激素
每个明显地分为皮质和髓质两部分。鸟类的肾上腺不形成明显的两层,髓质组织散在分布于皮质组织内。皮质和髓质无论在胚胎发生、形态结构和生理功能上,都是两个不同的内分泌腺。
一、肾上腺皮质
(一)肾上腺皮质激素
分为球状带、束状带和网状带3层。
球状带分泌的激素以醛固酮为主,主要参与调节体内水盐代谢,故称盐皮质激素(mineralocorticoid)。
束状带和网状带以分泌皮质醇为主,参与体内糖代谢的调节,故称糖皮质激素(glucocorticoid,glucocorticosteroid)。
网状带还分泌少量性激素(sex hormone),如脱氢异雄酮(dehydroepiandrosterone)和雌二醇(estradiol,E2)。
(二)盐皮质激素的生理作用
醛固酮的主要作用:是促进肾的远曲小管和集合管重吸收Na+、水,同时促进K+排出,称为“保钠排钾”作用。
(三)糖皮质激素的生理作用
1.对物质代谢的影响主要对糖、蛋白质、脂类和水盐代谢有影响。
(1)对糖代谢的作用是促进糖异生和抑制组织细胞对葡萄糖的利用,结果引起血糖浓度升高。
(2)对蛋白质代谢的作用既抑制蛋白质合成,又加速蛋白质分解,从而造成负氮平衡,其中对骨骼肌、骨、淋巴器官等的蛋白质代谢以促进分解为主,而对肝、胃、肠、泌尿和生殖等器官的蛋白质代谢则以抑制合成为主。
(3)对脂肪代谢的作用动员脂肪分解,使血中游离脂肪酸增多,并促进脂肪酸在肝内氧化供能。但由于在皮质醇作用下,血糖浓度升高,而刺激胰岛素分泌,胰岛素有强烈刺激脂肪合成的效应,所以皮质醇对脂肪代谢总的效应是促进脂肪合成。
2.参与应激反应当机体受到各种有害刺激,如创伤、手术、饥饿、寒冷等,血中ACTH浓度立即增加,糖皮质激素也相应增多。能引起ACTH与糖皮质激素分泌增加的各种刺激称为应激刺激,而产生的反应称为应激(stress)。
(四)皮质激素分泌的调节
1.糖皮质激素分泌的调节糖皮质激素的分泌受下丘脑-垂体-肾上腺皮质轴控制(图10-12)。垂体分泌的ACTH对束状带细胞类固醇激素的合成和分泌有直接的刺激作用,而ACTH分泌又受下丘脑CRH的控制。血中糖皮质激素的水平对其自身分泌可起反馈调节作用,这种反馈调节可作用于垂体、下丘脑或更高级脑水平上。
2.盐皮质激素分泌的调节醛固酮的分泌主要受肾素-血管紧张素系统的调节(详见第三章和第七章)。另外,血K+、血Na+浓度也可作用于球状带,影响醛固酮的分泌。
二、肾上腺髓质
(一)肾上腺髓质激素
肾上腺髓质分泌肾上腺素(epinephrine,E)和去甲肾上腺素(norepinephrine,NE)。它们都是儿茶酚的单胺类化合物,因此叫做儿茶酚胺。
(二)髓质激素的生理作用
NE主要调节循环系统的作用。
E主要调节体内的代谢变化。
当机体受到各种有害刺激,如创伤、饥饿、脱水、缺氧、寒冷等,这一系统将立即调动起来,儿茶酚胺(去甲肾上腺素、肾上腺素)的分泌量大增。儿茶酚胺作用于中枢神经系统,提高其兴奋性,使机体处于警觉状态;增强心血管系统和呼吸系统的活动,以利于应急时重要器官代谢的需要;促进糖和脂肪的分解代谢,以适应在应急情况下对能量的需要。
(三)髓质激素分泌的调节
1、反馈抑制:甲肾上腺素和多巴胺在髓质细胞内的量增加到一定程度时,可对其合成速度产生负反馈抑制;
2、交感神经:兴奋时引起NE释放,进而促进NE和E的合成。
3、ACTH和糖皮质激素:去垂体后,三种酶的活性下降,给以ACTH活性可恢复,给以糖皮质激素,也可使PNMT和DA羟化酶恢复活性。
第七节胰岛
胰腺的内分泌组织为胰岛,它散在于胰腺泡之间的各胰腺即是外分泌腺
内分泌腺
一、胰 岛 素
(一)胰岛素的化学
胰岛素是由胰岛的B细胞所分泌,由51个AA组成,分子量约为6000的小分子蛋白质,分为 A、B两条肽链。
(二)胰岛素的生理作用
胰岛素是调节三大营养物质代谢的重要激素,特别是对糖代谢的调节尤为重要。总的来说,胰岛素可促进三大代谢性营养物质以不同的形式在体内贮存,故胰岛素有“储存激素”之称。
1.对糖代谢血糖升高时,迅速引起胰岛素的释放,注射胰岛素在数分钟内血糖浓度即可降下来。
胰岛素可使全身各个组织加速摄取、贮存和利用G,特别是肌肉、肝、脂肪组织,结果使血糖水平下降。
2.对脂肪代谢胰岛素对脂肪代谢的作用主要有:
①促进肝细胞和脂肪细胞内脂肪酸的合成。
②促进葡萄糖进入肝细胞并使其转化为中性脂肪。
③抑制脂肪酶的活性,从而抑制贮存脂肪的分解。
④促进肝脏胆固醇的合成。
3.对蛋白质代谢胰岛素对蛋白质代谢的具体作用如下:
①促进氨基酸进入细胞。
②促进蛋白质的合成。
③促进RNA和DNA的合成。
④抑制组织蛋白质的分解,因胰岛素能稳定溶酶体,防止溶酶体蛋白水解酶的释放,从而抑制组织蛋白的分解。
(三)胰岛素分泌的调节
1.代谢物的作用血糖浓度升高时胰岛素的合成和分泌均增加,血糖浓度下降时则胰岛素的合成和分泌都相应减少。
血中氨基酸、脂肪酸和酮体等浓度的增加,也可使胰岛素分泌增多。
2.激素的作用
(1)多种胃肠激素(如胃泌素、促胰液素、胆囊收缩素和抑胃肽等)都在一定程度上具有刺激胰岛素分泌的作用。
(2)胰高血糖素通过旁分泌直接作用于B细胞使其分泌增多,也可通过血糖增高而间接地引起胰岛素的分泌。
(3)生长激素、皮质醇、孕酮和雌激素也能促进胰岛素的分泌。
3.神经调节支配胰岛的迷走神经兴奋时可通过M受体直接促进胰岛素的分泌,迷走神经还可通过刺激胃肠激素的释放,间接促进胰岛素的分泌。交感神经兴奋时,可通过α受体抑制胰岛素的分泌。
二、胰高血糖素
(一)胰高血糖素的化学
胰高血糖素是由胰岛A细胞所分泌。由29个氨基酸组成的直链多肽。
(二)胰高血糖素的生理作用
1.调节糖代谢胰高血糖素可促进糖原分解和葡萄糖的异生作用,而使血糖浓度升高。
2.调节蛋白质代谢胰高血糖素可促进组织蛋白质的分解和抑制蛋白质的合成,同时还可促进肝脏合成尿素。
3.调节脂肪代谢胰高血糖素可活化脂肪组织中的脂肪酶,促进脂肪的分解,使血浆中游离脂肪酸含量升高。
其它作用:促进胰岛素、生长激素、甲状旁腺素、降钙素和胆汁的分泌,抑制胃液的分泌。
(三)胰高血糖素分泌的调节
1.血糖浓度:。血糖浓度升高抑制胰高血糖素的分泌,而血糖浓度降低则促进其分泌。对维持饥饿条件下的血糖水平,保证脑的能量供应具有重要意义。
2.N调节:交感N兴奋,分泌增加;迷走N相反。
第八节其他内分泌腺和内分泌物质
一、胸腺
胸腺既是淋巴器官,又是具有内分泌功能的器官,它既能产生淋巴细胞又能合成和分泌胸腺激素(thymosin)。动物出生后胸腺继续生长,至性成熟时体积最大,以后逐渐退化,到了老年胸腺几乎被脂肪组织所代替。
胸腺激素是由胸腺髓部网状-上皮细胞合成和分泌的一组具有免疫活性的多肽类物质。目前已从胸腺抽取物中提纯出16种具有生物活性的多肽。根据这些多肽的结构和功能特点可将其分为三类。
第一类是促进细胞免疫应答因子(包括胸腺素、胸腺生成素、胸腺体液因子等)。这类因子能诱导淋巴干细胞成熟,转化为具有免疫性的T淋巴细胞,从而维持机体正常的免疫功能。第二类是抑制素,抑制素能降低T淋巴细胞的功能,抑制自身免疫功能。第三类是与免疫无关因子(包括低血糖因子、低血钙因子等)。
综上所述,胸腺激素的主要功能是刺激、诱导淋巴干细胞使其转变为T淋巴细胞,参与机体的细胞免疫。胸腺功能障碍可导致机体细胞免疫功能缺乏,易继发感染,甚至引起死亡。若胸腺功能亢进,常发生自身免疫性疾病,如重症肌无力,系统红斑狼疮等。
胸腺对类固醇激素比较敏感,性腺类固醇激素、肾上腺皮质类固醇激素对胸腺均具有一定的抑制作用。此外,甲状腺激素对胸腺有着促进作用。实验还证明胸腺与肾上腺皮质和性腺之间有交互的正反馈作用。
动物虽然在性成熟之后胸腺逐渐退化,但网状-上皮细胞仍然保持,胸腺的功能并没有丧失。成年动物如果由于各种原因(如放射损害、药物作用等)使淋巴细胞破坏和免疫活性细胞极度减少时,骨髓的淋巴干细胞进入胸腺,在网状-上皮细胞作用下成熟,转化为新的具有免疫活性的T淋巴细胞,然后又转移到淋巴结和脾脏,重新建立细胞免疫系统。
二、松果腺(松果体)
松果腺位于四叠体前丘之间的凹陷处,有一个柄与第三脑室背面相连,接受由颈上神经节发出的交感神经节后纤维的支配。松果腺一般在初情期以前已开始退化。松果腺所分泌的激素有两大类:①吲哚样类,如褪黑激素、5-甲氧基色醇等;②肽类,如8-精催产素、促性腺激素释放激素等。
褪黑素(melatonin,MLT),化学结构为N-乙醇-5-甲氧色胺,是由色氨酸转化而成。MLT具有抗生殖作用,MLT对下丘脑—垂体—性腺轴和下丘脑—垂体—甲状腺轴均有抑制作用,幼年时可以防止性早熟。如切除幼年动物的松果体,出现性早熟,性腺和甲状腺的重量增加,成年则可影响整个生殖过程。
MLT的释放有着明显的昼夜节律变化,即光照下分泌减少,黑暗时则分泌增加。
8-精催产素(AVT)具有抗利尿和催产作用,并对性腺活动具有很强的抑制作用。
三、胎盘及其激素
胎盘是由胎儿胎盘(尿囊绒毛膜)和母体胎盘(子宫内膜)共同构成。它是胎儿生长发育过程中与母体进行物质交换的临时性器官,同时它又是一个重要的内分泌器官。胎盘能分泌多种激素,如:雌激素、孕酮、松弛素、胎盘催乳素、胎盘促性腺激素、肾上腺皮质激素、生长激素,促性腺激素释放激素样物质、促甲状腺激素及β脑啡呔等。这里仅介绍2种胎盘促性腺激素。
(一)人绒毛膜促性腺激素
人绒毛膜促性腺激素(human chorionic gonadotrophin,HCG)是妊娠妇女胎盘胚泡滋养层细胞所分泌的一种促性腺激素,是一种糖蛋白,分子量约36000~40000。由于其分子量较小,可以从肾小球滤过,故可以在尿中测出。HCG具有垂体FSH和LH双重活性,而LH活性相对大于FSH活性。所以它的生理作用与LH相似,可促进排卵后黄体的形成。在妊娠早期可使黄体继续发育,在妊娠中期则代替卵巢合成雌激素和孕激素。此外HCG还能降低淋巴细胞的活力,防止母体产生对胎儿的排斥反应。根据其生理作用,畜牧业生产中广泛应用HCG来完成畜体的超数排卵和同期排卵,也可用于纠正排卵延迟或不排卵。
(二)孕马血清促性腺激素
孕马血清促性腺激素(pregnant mare serum gonadotropin,PMSG)是母马、母驴胎盘子宫内膜杯部的特殊细胞所产生的一种促性腺激素。母马妊娠40天开始分泌PMSG,并在55~75天的期间保持高峰分泌状态,以后分泌逐渐减少,到150天左右完全消失。
PMSG是一种酸性糖蛋白,分子量为70000,由于其分子量大,不能通过肾小球滤过,故只存在于血液中不存在于尿中。PMSG具有垂体FSH和LH的双重作用,而FSH活性相对大于LH的活性,所以它的生理作用与FSH相似,主要是促进卵泡的生长发育,也有一定的促排卵和促黄体生成的作用。畜牧业生产中被广泛用于实验动物和牛、羊、猪等的催情和超数排卵。
四、前列腺素
前列腺素(prostaglandin,PG)首先在动物的精液中发现。60年代之后发现哺乳动物许多器官系统均能分泌PG,而且几乎所有的组织和体液中均含有PG。
PG是由花生四烯酸转化而成的。根据环戊烷和脂肪酸侧链的不饱和程度和取代基的不同,可将天然前列腺素分为A、B、C、D、E、F、G、H和I等类型。PG的生理作用极为广泛和复杂,几乎每个器官和系统的活动都受到它的影响。同一种前列腺素对不同组织有不同的作用;而同一种组织对不同的前列腺素发生的反应也很不相同。
PG的半衰期仅1~2分钟经肺和肝迅速降解,血中浓度很低,因此不属于循环激素,而是在局部组织产生和释放,通过旁分泌的形式调节局部生理机能的一组局部激素。
在畜牧兽医生产实践中,利用PGF2α和PGE2对黄体的溶解作用来控制动物的发情(引起同期发情),以利于实施人工授精。同时亦利用PGE2或PGF2α能刺激子宫肌的收缩,而达到催产的目的。
五、细胞生长、分化的有关因子
动物体内存在着各种各样促进细胞生长和分化的因子,它们都属于多肽活性物质。这些活性物质的受体大多数均具有依赖酪氨酸蛋白激酶的活性,即其本身就是蛋白激酶,可发生自身磷酸化,然后促进细胞的生长和分化。这些因子在体内组成复杂的“生长因子”系统。近来还发现细胞中还存在生长抑制因子,称为抑制素(inhibin)。
(一)上皮细胞生长因子
上皮细胞生长因子(epidermal growth factor, EGF)存在于唾液腺中,是由53个氨基酸残基组成的单链多肽,分子量为6000。EGF的受体广泛分布于不同细胞的膜上,是一种依赖酪氨酸的蛋白激酶。由于EGF具有刺激上皮生长的作用,故命名为细胞生长因子。在胚胎生长发育过程中可能起着促进上皮细胞增殖和分化的作用。
(二)胰岛素样生长因子(insulin-like growth factor, IGF)
实验证明生长素对软骨组织生长并无直接作用。生长素能促进肝、肾等组织产生生长素介质(SM),SM进而促进细胞的生长。目前已分离出两种生长介素,其化学结构与胰岛素原相似,故称为胰岛素样生长因子(IGF),IGF有IGF-Ⅰ和IGF-Ⅱ(IGF-Ⅰ即生长介素C,IGF-Ⅱ即生长介素A)。IGF是由70个氨基酸组成的单链多肽,IGF-Ⅱ比IGF-I少3个氨基酸。它们的受体相似,也具有依赖酪氨酸的蛋白激酶活性,可发生自身磷酸化从而刺激细胞的生长和增殖。
另外,还有促纤维细胞生长的成纤维细胞生长因子(fibroblast growth factor, FGF);促进细胞有丝分裂的血小板衍生因子(platelet-derived growth factor, PDGF)和促进神经细胞生长的神经生长因子(nerve growth factor, NGF)等。
第十章 生殖生理
第一节Introduction
Sex来自拉丁词Sexus,它的意思为分化,分开,切断的意思。
一、性征、性决定和性分化
区别雌雄的特征有第一性征和第二性征之分。
第一性征是产生配子的性腺,如睾丸、卵巢。
第二性征包括形态、机能和心理三方面。
生殖器第二性征(附性器官)是运送配子的管道和相连的腺体。
男:附睾、输精管、尿道、阴茎、前列腺、精囊及尿道球腺
女:输卵管、子宫、阴道、生殖道粘膜中的腺体
生殖器外第二性特征:是区别男女性别的特征,这些特征自青春发育期逐渐突出,成年后最为明显,性征是性决定和性分化的结果,生物学上性的决定和分化是由染色体的性别、性腺和生殖器的性别、身体的性别、生后的养育和社会心理学上的性别等多个环节和步骤相继发育,交互影响造成的。
二、生殖过程
新生个体的产生是两性性细胞结合后,在母体的子宫经发育、生长、最后娩出的过程。
三、性成熟和体成熟
1、性成熟(sexual maturity):哺乳动物生长发育到一定时期,生殖器官基本发育完全,并且具备繁殖能力,这一时期叫做性成熟。
初情期:母畜第一次发情或第一次排卵的时期。此期发情与发情周期的周期不正常或不规律,经过一定时期达到性成熟,开始排成熟的卵子。
如何确定雄性初情期?一次射精50000000个精子,其中10%以上是具有活力,即具有受精能力。
性成熟的年龄随动物性别、气候、营养和管理而有所不同,一般讲:小动物比大动物性成熟早,如牛8-12个月,家兔4-5个月,羊4-8个月,臭虫:一晚72代,母畜比公畜早,气温高的地区比气温低的地区家畜早。
2、体成熟:动物的生长基本结束,并具有成年动物所固有的形态和结构特点,称为体成熟。
体成熟比性成熟晚,性成熟后身体仍在发育、生长,到一定时间后才达到体成熟,因此家畜开始配种的年龄要比性成熟晚些。
四、性季节(繁殖季节)
家畜的繁殖有季节性,这是长期自然选择的结果,因为在未驯养前,处在原始的自然调节下,只有那些再全年中比较良好的环境条件下产仔的,才能保证其新生的幼仔能够活下来,如马,发情在春季,妊娠11个月,则分娩在春季,绵羊的繁殖季节在秋季,妊娠5个月,则分娩季节也为春季。
牛、猪、兔—终年多次发情,马、羊、驴在一定季节呈多次发情。
第二节雄性生殖生理
一、睾丸的功能
睾丸是雄性动物的主性器官,它具有产生精子和分泌雄性激素的两大功能。
(一)睾丸的生精作用
睾丸由曲精细管与间质细胞组成,曲精细管上皮有两种C:精原C(精子)与支持C(营养C,起营养和支持作用)。
精子生成后储存在附睾内,射精释放,不射精,经一定时间后将衰老、死亡并吸收。
支持C与支持C之间有紧密连接( tight junction),而其底部又与基膜紧贴,这一特殊结构使体液成分难以直接进入曲精细管管腔,形成“血睾屏障”。“血睾屏障”可以防止生精细胞的抗原性物质进入血液循环而引起自身过敏性反应。
精子的生成需要适宜的温度(阴囊内较腹腔低4-7度)
(二)睾丸的内分泌功能
1、睾丸激素睾丸的间质细胞可产生雄性激素(androgen)和少量雌激素,化学本质是类固醇,作用最强的是睾丸间质C所分泌的睾丸酮—睾酮。
2、雄性激素的生理作用:
①维持生精。睾酮进入曲细精管转变为双氢睾酮,与ABP结合,促进生精。曲细精管微环境中高浓度的雄激素是维持生精的必要条件。②促进雄性生殖器官的生长发育及副性征的出现,维持正常的性欲和性反射。睾酮主要刺激内生殖器(曲细精管、输精管、附睾、精囊、射精管等)的生长发育,而双氢睾酮则促进外生殖器(尿道、阴茎、前列腺等)的生长发育。③促进蛋白质的合成,特别是促进肌肉和生殖器官的蛋白质合成,从而使尿氮排出减少,呈现正氮平衡。④促进骨的生长和钙磷沉积。⑤促进红细胞的生成(睾酮可促进红细胞生成素的生成)。
二、睾丸活动的调节
睾丸的活动受到下丘脑—腺垂体—性腺(睾丸)轴及其靶激素的反馈性调节。
(1)在内外环境因素的作用下,下丘脑可释放GnRH,GnRH通过垂体门脉作用于腺垂体,促进腺垂体分泌FSH和LH。FSH和LH都能控制精子的生成。
(2)雄激素在血浆中达到一定浓度时,可反馈性抑制GnRH和FSH、LH的分泌,从而使睾酮的分泌量维持在一定的水平上。
(3)支持细胞所分泌的抑制素(inhibin)是蛋白激素,由α、β两个亚单位合成,分子量为31000-32000,它对FSH的释放具有很强的负反馈抑制作用,而同样生理剂量的抑制素对LH分泌却无明显影响。
三、附睾及副性腺的主要功能
(一)附睾的主要生理功能
附睾为细长管道,叫附睾管,功能主要为对精子的转运、浓缩、成熟和储存。
(二)副性腺的主要生理功能
指尿道球腺、前列腺和精囊腺3种。它们的分泌物共同组成精液的液体部分(或叫精液)内含果糖蛋白。
1.尿道球腺分泌透明粘液,在射精前排出,呈碱性,主要冲洗尿道和中和阴道内酸性物质的作用。
2.前列腺分泌物较稀,不透明,有特殊臭味,含阮较多,呈碱性,可中和阴道内酸性物,并吸收精子排出的CO2以利于精子的活动。
3.精囊腺分泌量大,白色胶状粘液,进入雌性阴道中可凝固成栓,防止精液倒流,有利于受精。
四、精液
(一)精子
精子是在睾丸的曲细精管内产生、贮存于附睾并由附睾排出的。它是雄性动物的生殖细胞,带有父本遗传信息。
精子由头和尾两部分组成。头部包括细胞膜和顶体等,能进入卵细胞并与卵细胞结合,而尾部则是精子运动的部分。
精子运动的动力:向流性:逆流游动
向触性:向异物
向比性:向化学物质
(二)精清
精清的主要生理作用有:①稀释精子,便于精子的运行和输入雌性动物的生殖道。②提供精子运动和存活的适宜环境。③提供精子活动的能源(精清含有果糖、山梨醇和甘油磷酸胆碱等能源物质)。④保护精子、防止氧化剂对精子的损害(精清中的巯基组氨酸三甲基钠盐具有抗氧化作用),和防止精子的凝集(精清中含有抗精子凝集素)。⑤精清中的PG能刺激雌性生殖道的运动,有利于精子的运行。⑥有些动物的精清能在雌性生殖道内凝固成栓塞,防止精液倒流。
第三节 雌性生殖生理
雌性生殖包括卵细胞的形成、排卵、受精、妊娠和分娩等生殖过程。
一、卵巢的功能
(一)卵巢的生卵作用
雌性生殖过程有明显的周期性,卵子的生成与性腺的分泌活动是有节律进行的,这与体内激素调节有关,这种生殖周期,在灵长类以外的哺乳动物称动情周期,灵长类称月经周期。
母畜达到性成熟的主要生理特征是卵C在卵巢中的成熟和排出。
卵子的发生和卵泡的发育过程如下:
性C→卵原C→初级卵母C→ 次级卵母C→ 卵母C→ 初级卵泡→次级卵泡→ 卵子→成熟卵泡→ 破裂排卵
卵子的结构:包括放射冠、透明带、卵黄膜及卵黄。
牛、马、蹄类、灵长类动物每次发情只排一个卵子;猪、山羊、犬、兔可排出两个以上卵子。
成熟卵泡破裂后,卵泡的外壁因压力减小而塌陷,卵泡腔内充满着由卵泡膜血管破裂时流出的血液,以后卵泡上皮细胞又逐渐形成新的C层,代替血凝块,并在细胞的原生质内积蓄黄色颗粒,使破裂的卵泡形成黄体。黄体存在的时间要看是否受精而定,若是卵子已受精,黄体就继续生长,这时叫妊娠黄体,直到妊娠末期才逐渐萎缩,如未妊娠,黄体不久就萎缩退化,最后形成一个白色物叫白体。
黄体可以看作是一个内分泌器官,LH通过cAMP使黄体分泌大量孕酮和少量雌激素,一般于排卵后7-8天,黄体发展到顶峰,若没受精,黄体则退化。
卵巢活动的周期性变化:
卵泡生长发育成熟→排卵→ 黄体生长 →退化→下一周期
每一周期包括以下两个期:卵泡期:
卵泡由生长、发育到排出的时期
黄体期:黄体生长发育到退化的时期
(二)卵巢的内分泌功能
卵巢除产卵外,还能分泌性激素,有雌激素(estrogen, E)、孕激素(progestogen)及少量的雄激素。妊娠期间还可分泌一种使耻骨韧带松弛的松弛素。
人的卵巢只能合成雌二醇和雌酮。
1.雌激素及其生理作用体内雌激素包括雌二醇(E2)、雌酮及雌三醇,化学结构亦为类固醇。雌二醇是由卵巢的卵泡内膜C分泌的,效力最强,另外两种是雌二醇的代谢产物,怀孕期间,胎盘产生大量雌三醇。
雌二醇的生理作用:
(1)刺激附性器官的生长:使子宫内膜增厚,增加血流和C活动,提高子宫对催产素的反应,为怀孕做准备。
(2)促进阴道上皮的增生和角化,增强抵抗力。
(3)刺激输卵管的生长和运动。
(4)促进乳腺导管系统得生长发育。
(5)与雌体形发育有关。
(6)促进水、Na、Ca、N和P的潴留,对骨代谢有影响。
(7)促进发情行为和表现。
(8)大剂量能抑制FSH的分泌,而刺激LH分泌。
2.孕激素及其生理作用孕激素主要是由黄体和胎盘所分泌的,类固醇,以孕酮作用最强,它在尿中的代谢产物是无活性的孕二醇;马和绵羊由胎盘产生孕激素,黄体消失后不会发生流产;猪、山羊、兔的黄体是产生孕激素的唯一来源,因此,妊娠阶段切除卵巢,会引起流产。
孕H通常要在雌H作用的基础上才能发挥它的作用。
二、性周期(发情周期)
母畜性成熟后,卵巢中就规律性的出现乱泡成熟和排卵过程,哺乳动物的排卵是周期性发生的,伴随每次排卵,母畜的机体特别是生殖器官,发生一系列的形态和生理性变化。
性周期:指从一次发情开始到下次发情之前,或由一次排卵到下次排卵的结个间隔时间。
马、驴、牛、猪→多周期动物,狗及多种野生动物→少周期动物;绵羊在每一季节内出现几个彼此相继的性周期→长乏情期
性周期的分期:
除灵长类动物以外,所有哺乳动物的性周期又叫发情周期(Estrus cycle),根据母畜生殖器官所发生的变化,一般可分为发情前期(proestrus)、发情期(estrus)、发情后期(metaestrus)、休情期(diestrus)四个时期。
三卵巢活动的调节
内、外环境的变化可通过下丘脑—腺垂体—性腺(卵巢)轴及靶腺激素的反馈作用来调节卵巢的活动。
(1)在内外环境因素的作用下,下丘脑释放GnRH,作用于腺垂体使其释放FSH和LH作用于卵巢。
(2)FSH与类胰岛素生长因子和上皮因子等可促进卵泡的生长发育并使其分泌雌激素,同时能使颗粒细胞产生芳香化酶,将内膜细胞产生的雄激素转化为雌激素。而血中雌激素达到一定浓度时,又可对GnRH和FSH的分泌产生负反馈作用。LH作用于内膜细胞,使胆固醇转变为雄激素。
(3)卵巢中的雌激素又可通过局部正反馈作用,增加卵泡对LH和FSH的敏感性,在FSH分泌减少的情况下,继续促进卵泡的生长发育。在排卵前夕高浓度的雌激素又对GnRH和LH的分泌产生正反馈作用。
(4)卵泡的颗粒细胞能分泌抑制素,对FSH的分泌产生抑制作用。
(5)PRL具有维持黄体的作用。
通过下丘脑—垂体—卵巢轴对卵巢活动的调节及卵巢内的自身调节过程,既保证血液中雌激素和孕激素浓度的水平,又保证性周期不同时期对雌激素和孕激素的需要,从而维持雌性动物的正常生殖过程。
四、附性器官的功能
(一)输卵管的功能
输卵管的主要生理作用如下:
1.接纳卵巢排出的卵子卵巢排出的卵子,一般均被纳入输卵管的伞端。
2.卵子和精子的转运作用在卵巢激素的作用下,可促使输卵管上皮纤毛和管壁肌发生有规律的蠕动,而使精子和卵子分别向着输卵管上1/3的壶腹部转运。
3.输卵管还是精子获能和受精的地点(详见受精)。
4.是受精卵卵裂和早期胚胎发育的场所输卵管分泌细胞分泌的液体可提供受精卵卵裂、胚胎早期发育的营养,有利于受精卵向子宫方向转运。
(二)子宫的机能
子宫是胚胎发育的场所,妊娠期所形成的胎盘是重要的内分泌器官。子宫的主要生理作用如下:
1.子宫肌的运动对生殖机能的影响发情期在卵巢激素和交配等因素的作用下,子宫肌发生节律性的收缩,可促进精子向输卵管方向移动,有利于受精;妊娠期在孕激素的作用下,子宫肌运动减弱,处于相对静止,有利于胎儿的生长发育;分娩时,在神经体液的调节下,子宫肌发生强力收缩,促进胎儿排出。
2.提供胎儿生长发育所需的各种物质和环境胎儿在生长发育过程中所需的所有营养物质及其代谢产物的排出,均是通过胎盘而实现的,胎盘是母体子宫组织和胚胎组织共同构成的临时性器官。
3.子宫能分泌前列腺素前列腺素可通过子宫-卵巢的局部循环而引起黄体溶解。
4.子宫颈分泌的粘液的作用发情期子宫颈分泌较为稀薄的粘液而有利于精子的通过;在妊娠期,其分泌物粘稠,闭塞子宫颈,可以防止感染物进入子宫。
5.子宫的内分泌功能(见内分泌章)。
第四节 生殖过程
包括受精、妊娠和分娩
一 受精
受精(fertilization)是指两性配子(卵子和精子)结合而形成一个新细胞—合子的复杂生理过程。它包括精子和卵子的运行、精子的获能作用、精子和卵子的相遇及顶体反应、精子进入卵细胞及合子的形成和透明带反应等重要生理过程。
1.精子的运行
精子在母畜生殖道内由射精部位到受精部位的运动过程。
精子运行的能力
来自本身的运动(次)
借助母畜输卵管的收缩和蠕动(主)
精子进入母畜生殖道之后,须经过一定变化后才能具有进入透明带和使卵子受精的能力,这一变化过程叫精子的受精获能过程(或受精获能作用),获能时间大约在子宫或输卵管内停留2-4小时。
2.卵子的运行
排卵时,卵子随卵泡液被纳入输卵管伞部,并借助伞部上皮C纤毛的颤动和平滑肌的收缩很快进入输卵管,卵子在输卵管内运行时发生着变化,马、狗和狐的卵子排出时是初级卵母细胞,在输卵管中成熟,绵羊、牛和猪排卵时是次级卵母细胞。
次级卵母C包括一个核,核心周围是卵黄,卵黄的边界是卵黄膜,卵黄膜被一层厚而透明的膜保围着,这层膜叫透明带。
卵子保持受精能力的时间:即卵子运行到输卵管峡部以前的时间。
3.受精过程
(1)精子与卵子相遇:精子与卵子在输卵管壶腹相遇而受精,射精9亿个精子 →几千个到壶腹→1个精子进入卵子而受精
(2)精子进入卵子:精卵相遇,精子顶体释放出透明质酸酶,溶解卵子周围的放射冠,穿过透明带之后,卵子产生受精素与精子起特异性反应,使精子固定在透明带某一点上,继而精子又释放顶体素(蛋白水解酶),使精子突破透明带,而达卵黄膜,精子失去顶体,头进入卵黄膜→发育
(3)原核形成和配子组合
精子进入卵子后,脱掉尾巴,头部膨大,细胞核形成雄性原核,卵子的核形成雌性原核,两个原核接近,核膜消失,各自形成染色体,进行组合,完成受精的全过程,接着发生第一次卵裂。
二、妊娠(pregnancy)
受精卵在母体子宫内生长发育为成熟的胎儿的过程。
- 卵裂和胚泡种植:
受精卵(合子)沿输卵管向子宫移动的同时,进行细胞分裂,叫卵裂。约3天,变成16-32个细胞的桑椹胚,约4天,桑椹胚即进入子宫,继续分裂,体积扩大,形成中央含有少量液体的空腔,叫胚泡。在胚泡周围形成一层滋养层,供给胚泡迅速增殖所需的营养,其后胚泡逐渐埋入子宫内膜而被固定,叫种植。种植后胚泡继续生长,由母体供给养料和排出代谢产物。
- 胎盘形成与胎儿发育
种植后的胚泡滋养层迅速向外增生,形成含胚泡血管组织的绒毛,与此同时,子宫内膜与胚泡相接的粘膜增生,形成覆盖胚胎的脱膜,绒毛深入脱膜内构成胎盘,从此胚胎在胎盘内发育成胎儿。
- 母体妊娠时的变化
妊娠黄体分泌大量孕酮,起安胎作用。在雌激素的协同作用下,袭击乳腺腺泡生长,使乳腺发育完全,准备泌乳。
随胎儿增大,引起消化、呼吸、循环、排泄等一系列变化,如胸式呼吸,呼吸浅而快,血沉加快,心脏负担增加,排尿、粪次数增加,代谢旺盛,前期食欲旺盛。
- 妊娠期间的发情
母畜发情周期一般由妊娠开始而中断,但有的妊娠母畜还可能出现发情,叫妊娠期发情。
- 假妊娠
母畜发情排卵后,如卵子并没有受精,而黄体继续存在,经一定时间后,出现乳腺发育、泌乳、做窝等妊娠征候,这一现象叫假妊娠。
三、分娩
分娩时成熟的胎儿自子宫排出母体的过程。通常分三期:
开口期:子宫节律收缩,子宫颈扩大。
娩出胎儿期:子宫更为频繁而持久收缩,腹隔肌收缩,内压升高。
胎衣排出期:胎儿出,经短时间间歇,子宫又收缩排出胎衣。
可见,胎儿从子宫中娩出的动力是靠子宫和腹壁肌的收缩来实现的。
第十一章 泌乳生理
母畜分娩后,以乳汁哺育其幼畜的过程,叫授乳。泌乳包括乳的分泌和排出两个独立又相互联系的过程。
泌乳期=哺乳期 猪:60天黄牛、水牛:90-120天,人工选育延长泌乳期300天。
第一节乳腺的结构
乳腺由皮肤腺体衍生而来,牛 2对,马、羊 1对,猪 5-8对
乳房内主要有两种组织:一种是由乳腺腺泡和导管系统构成的腺体组织或实质,另一种是由结缔组织和脂肪组织构成的间质,它保护和支持腺组织。
腺泡(基本单位) → 细小乳导管 → 中等乳导管 → 粗大乳导管 → 乳池(系乳房下部及乳头内贮藏乳汁的较大腔道,又叫乳槽) → 乳头管开口
牛每一乳腺各有一个乳头孔,马2个,猪2-3个,狗十几个,人15-25个乳头孔。
乳腺的血液供应十分丰富,每一腺泡均保围着毛细血管,为乳腺生乳提供应营养物质和氧。
第二节乳腺的发育及其调节
1、发育
2、乳腺发育的调节
(1)乳腺发育中内分泌腺的控制:乳腺发育是多种内分泌腺综合作用的结果,卵巢分泌的雌H和黄体分泌的孕酮参与调节乳腺的发育
三重切除试验证明:
性成熟前切除卵巢:乳腺发育不全,反之给未性成熟的母畜周期性的使用雌H时,可使乳腺发育。雌H:孕H=1:1000的情况下,牛乳腺能获得很好的发育;切除兔和大白鼠的脑垂体、卵巢和肾上腺的研究表明:手术后若适当注射雌H + 孕H + GH + PRL + 肾上腺皮质H,可刺激萎缩的乳腺导管系统充分发育达妊娠后期的程度,从而促进乳腺小叶—腺泡的发育。
(2)乳腺发育的神经调节
刺激乳腺的感受器 → 中枢神经系统 → 丘脑下部垂体系统 → 传出神经 → 控制乳腺发育按摩
神经系统还具有营养作用切断乳腺N乳腺发育停止
第三节乳的分泌
乳腺组织的分泌C,从血液中摄取营养物质生成乳汁后,分泌入腺泡腔内,这一过程叫做乳的分泌(milk secretion)。
1、乳汁的成分及性质
初乳:在分娩期或分娩后最初3~5天内,乳腺产生的乳叫初乳。初乳较粘稠,浅黄,初乳干物质含量高,含有丰富的球蛋白和白蛋白、酶、Vit及溶菌素,含较多的无机盐,Mg盐有轻泻作用。
常乳
初乳期过后,乳腺所分泌的乳汁,叫做常乳。
含 a水b蛋白质:以酪Pr为主,其次是白Pr和球Pr。c脂肪:是油酸、软脂酸及其它低分子脂肪酸的甘油三酯。d糖:过氧化氢酶、脱氢酶、水解酶、过氧化物酶。e Vit:A、B、C、D、色素、抗毒素。f 无机盐:氯化物、磷酸盐、硫酸盐
2、乳的生成过程
乳的生成过程是乳腺腺泡和细小乳导管的分泌上皮细胞内进行的。包括一系列新物质的合成绩复杂的选择性吸收过程。乳中的球蛋白、酶、H、Vit和无机盐等均由血液进入乳中,是乳腺分泌上皮对血浆选择性吸收和浓缩的结果。
乳的合成
血液中的氨基酸合成蛋白质。葡萄糖转变为乳糖。乳脂几乎完全呈甘油三酯状态,它在上皮C的颗粒性内质网中形成脂肪小球。
3、乳的分泌过程
乳腺分泌C内合成的或选择性吸收的乳组分,它的分泌过程包括两步:第一步是乳组分从其合成部位经C浆到达顶端C膜,;第二步越过C膜进入腺泡腔中,乳脂、乳蛋白、乳糖及K、Na的分泌建书P331。
4、乳分泌的调节
泌乳期间乳的分泌包括引起泌乳和维持泌乳两个过程,主要通过N—H途径来进行调节。
脑垂体前叶的催乳H在妊娠期间被胎盘和卵巢分泌的大量的雌H和孕酮所抑制,分娩后,孕酮水平突然下降,结果PRL迅速释放,对乳的生成产生强烈作用,引起泌乳。以后低水平的雌H及分娩后高浓度的肾上腺皮质H均加强PRL和GH的泌乳作用。
泌乳期间垂体分泌PRL是一种反射活动,主要是由哺乳或挤乳对乳房的刺激。
甲状腺和肾上腺皮质通过支配乳腺的分泌N纤维和营养NF影响乳的生成过程。
4、乳的排出
乳腺泡的上皮细胞生成的乳汁,连续地分泌到腺泡腔内。当腺泡腔和细小乳导管充满乳汁时,则腺泡周围的肌上皮细胞和导管系统的平滑肌反射性收缩,将乳汁转移入乳导管和乳池内。乳腺的全部腺泡腔、导管、乳池构成蓄积乳的容纳系统。
当哺乳或挤乳时,引起乳房容纳系统紧张度改变,使贮积在腺泡和乳导管系统内的乳汁迅速流向乳池,这一过程叫做排乳(milk excretion)。
乳池乳:乳头括约肌开放时,乳池内的乳借助本身重力作用而排出的乳。牛一般占1/3—1/2左右。
反射乳(reflex milk):由排乳反射从乳腺腺泡及乳导管所获得的乳叫反射乳。占总产量的1/2—2/3。
排乳的神经—体液调节
排乳过程是由条件和非条件反射组成的复杂的反射活动。
排乳反射的传出途径有:(1)传出NF存在于精索外N和交感N中,直接支配乳腺平滑肌的活动。
(2)通过下丘脑—垂体的N-体液途径来调节排乳活动。
交感N兴奋(纯粹的N反射)→第二种反射(N-体液排乳反射) →催产素→大脑皮质→排乳的抑制反射:中枢阻止N垂体释放催产素,外周交感N使肾上腺髓质释放肾上腺素。
生理学的发展(Physiology history)
生理学是生物科学(biological science)的一个分支,是研究生物机体的各种生命现象。
physiology的含义,19世纪法国著名的生理学家Clande Bernard描述:“生理学是关于生命的科学”。定义:研究人体及动物机体的各种机能(function)及活动规律的科学。各种机能是指—人体和动物的整体及其各组成系统。器官和细胞所表现的各种生理活动,如,血液循环、呼吸、消化、排泄、生殖、肌肉运动和N活动等生理现象产生的过程和机制(mechanism),以及机体内外环境因素对这些生理机能的影响。如:泌尿、、、、、,人体和动物机动物机体只有在存活时,才表现这些复杂的生理活动,所以生理学研究的基本原则是以活体作为研究对象,生理学泛指人体及动物生理学,是研究人体及动物机体的生命活动规律的科学。形态学研究的一般都是尸体(静态无时间变化),而生理学研究的必须是活体(动态有时间变化)。
就生理学而言,研究内容十分广泛,因生物门类多,动物有1百万种,植物30多万种,微生物10多万,它们的结构与功能变化极大,因此分支多,由此形成很多分支学科,并且分支越来越细。如,细菌生理学、植物生理学、动物生理学,它又可分为:哺乳、鸟类、昆虫、鱼类。哺乳类分人体和家畜。人体与医学有直接关系,人体生理学研究比较深入,但由于在人体实验受限,所以用接近人体的哺乳动物,如:因此,动物生理学发展较突出。
生理学和形态学(morphology)在十九世纪后半期划分开来,不久又从生理学分化出生物化学,之后又分化出生物物理学,
生理学(physiology)这一词人们并不陌生,因为他与医学有着极其密切的关系,他的形成与发展是人们在长期的生产劳动和生活实践中不断地和疾病作斗争,逐渐积累了许多有关人体机能的知识。
一)古代和中世纪的生理学知识:
古代希腊名医Galen(129-199)对人体生理学体系曾提出“三元气”学说,认为生命的基本要素是元气。从肝中产生了带有自然元气的血液,流到心脏后变为生命元气,这些元流到脑基底部的颈动脉的末端分枝,变成动物元气,然后这些微妙的液体通过中空的管状N流到全身,他认为N是空心的。动脉内充满了气体。因他解剖的尸体中A中血液都已流入V,虽然Galen的这种假设是没有实验根据的,但却一直流传到十七世纪,这是由于在欧洲的黑暗时期和中世纪,解剖人体是被禁止的,直到十五世纪后半期“文艺复兴”时期人体解剖学才得到发展,直到十六世纪比利时学者Vesalius(1514-1564)认真从事尸体解剖从而写出了名著《人体构造》纠正了Galen解剖学中许多关于人体结构成错误识法。哈维在意大利padua大学医学院学习并毕业于学校,致力于用活体解剖的方法,从实际观察中研究心脏的活动,1628发表了《动物心脏及血液运动的解剖学研究》从而论证了他的新学说。
1.由于心脏的活动,血液不断地由从腔V输送到A,其量之大是不可能由吸收的营养物中提供,而且全部血液是以很快的速度通过心脏,通过计算,只半小时,心脏射入大动脉的血量就相当于或超过全身的血量。
2.V从身体各部分把血液不断地送到心脏,利用手臂的皮下V演示了V中的血流方向与辩膜关系。
说明在V中,由于辩膜的作用使血液在心脏流动。
《动物心脏及血液运动的解剖学研究》发表以后,才推翻了Galen的许多不符合事实的,创立了解剖生理学派。
Physiology一词是在16世纪Jean Fernel开始用 Aristotle(384-322 BC, Greek philophers)提出的physiologia一词来称呼研究人体结构与功能的这一部分医学,到十七世纪,当时仅是医学中的一章,真正生理学作为一门独立的学科是在英国医生William Harvey发现了血液循环之后。
二、生理学的发展:
1628年英国名医William Harvey(1578-1657)出版了《心血运动论》一书,这是历史上首次出现有确凿证据的生理学著作,从此生理学真正作为一门实验性学科。从而奠定了现代实验生理学的基础。他的结论是在几种动物身上应用活体解剖法通过多次实验获得的。因此,哈维的历史功绩不仅在于血液循环的发现,而是通过发现血液循环把实验方法引入了生物学,因此后人把1628哈维发现血液循环作为生理学成为实验科学的里程碑。在十七、十八世纪,显微镜的发明与化学物理学的迅速进步都给生理学的发展创造了良好条件,1661年意大利组织学家,马尔比基(M Malpigi1628-1694)发现了动脉与V之间的毛细血管。
应用显微镜发现了毛细血管的结构后,才真正完成了血循的线路。
法国哲学家R Descartes(笛卡尔)1596-1650依异物碰到角膜即引起眨眼等类现象,首次提出了反射的概念,Reflex.认为动物的活动都是对于外界一定刺激的反映。这为以后NS生理学发展指出了方向。
俄国学者罗蒙诺索夫(1711-1765)首次提出物质守恒与能量守恒及转化定律,以及后来法国化学家Lavoisier 1743-1794关于燃烧和呼吸原理的阐明,给机体新陈代谢的研究打下了基础,到了十九世纪,随着其它自然科学的迅速发展,积累了大量有关器官生理机能的知识如:
德国学者T Miiller(1801-1858)关于器官的研究。
E.Du.BoisReymond 1818—1896关于肌肉的研究。
K.ludwig关于循环、排泄的研究。
法国学者Claude Bernard 1813—1878关于代谢和机体内环境的研究
十九世纪末至二十世纪初,在分析生理学理论与实验方面影响最大的是英国学C S Sherrington(1857-1952)关于脊髓反射机制的研究,在1906年出版的《NS的整合动作》一书,给了反射理论以丰富的客观实验证据,提供了NS活动的一些基本规律,1863俄国学者谢切诺夫(1829-1905)发表了名著《脑的反射》将反射概念应用于脑的活动,认为脑的活动的本质是反射活动,后来,巴甫活夫1849—1936发展了生理外科和慢性手术方法研究消化的机能,在本世纪初,他集中地研究了大脑皮质生理学创出了高级N活动学说,总结出一系列关于高级N活动的基本理论。条件反射和非条件反射。
由于科学技术的进步及化学、物理学和数学的发展二十世纪形成了许多新的科学分支,本世纪二十年代初形成了生化,五十年代建立了生物物理学,六十年代出现了生物数学。
目前发展的趋势,一方面向分子生理学发展,另一方面逐渐打破学科界限,对机体的机能进行全面综合性的研究形成许多边缘学科,如N生物学E把组织学家,胚胎学家、解剖学家、生理学家、药理学家、生化学家、生物物理学家、心理学家以及生态学家,遗传学家、教育学家、人类学家、从各角度汇集起来,打破了传统学科的框架,共同解决脑的功能。
科研:鸡蛋生物学的研究,生理、电子、机械。
- 绪论
一、生理学的研究对象
动物生理学研究动物体,(包括人体)的各种机能及其活动规律的科学,研究这些机能在动物体内如何协调,如何密切配合形成一个统一的整体,动物体又如何与外界环境相适应等。
对复杂生命现象可分别进行不同层次的研究,即对生命现象发生发展规律,从不同角度,不同水平去研究,现代生理学大体上以三个不同水平进行研究。
1、从整体与环境之间,以及体内各器官系统生理活动之间的相互关系去研究,也就是研究整体对环境变化的反应与适应,这包括研究自然环境的变化,如气温、光照、缺氧等环境因素对生理活动的影响,以及机体对这些变动因素的适应能力和限度,也研究机体各系统生理活动是如何通过复杂的调节系统进行相互配合和适应不断变化的环境条件,如①寒冷,机体对寒冷的刺激,机体内发生一系列变化
②高山缺氧。
2、从器官和系统水平去研究,因为整体的生命活动是以各器官的生理活动协调为基础的,要认识整体生命活动规律,首先应该认识各器官的生理活动,也就是各器官有哪些特殊的生理活动,这些活动是如何发生发展的,受哪些因素的影响和制约,在整体生命活动中起什么作用,这类研究是以器官为对象的。这个范畴的研究属于器官水平的研究,对各器官生理活动的研究器官生理学,如:循环生理。
3、从分子和细胞水平去研究,因为器官是由不同生理特性的细胞构成,而且细胞的生理特性又决定于这些细胞组成物质的物理化学和生物化学的变化过程,细胞的组成物质主要是proteins, lipids、Nucleic Acids Carbohydrates.这些统称为Biomolecule,biological molecule。细胞和水平的研究主要研究细胞内亚微结构的机能,以及各种生物分子的理化变化,如心肌收缩,要研究心肌细胞膜、内质肉和肌原纤维等亚微结构的机能活动以及心细胞的收缩蛋白质与有关离子和酶相互作用的理化过程,这类研究为细胞生理学—即研究细胞的结构和功能。
例子:(目的进一步解释)
如:在可怕的事件发生时,紧张或运动时,从整体看,机体处于紧张状态,即应激状态,表现为瞳孔扩张(大),中枢NS反应快(迅速),心率上升,血压上升,呼吸加强,血糖上升,体内激素发生变化。机体为什么发生这样的变化,机体内各器官系统是如何协调的,这就要以整体不平去研究。
进一步分析心脏是怎样射血的,心率快慢受哪些因素影响,等等,这需从器官和系统水平去研究。
心肌为什么能收缩,解释这一解,必须研究心肌细胞膜,内质网,如肌原F等亚微结构的机能活动,以及心肌细胞中蛋白质等生物分子与有关离子运动的物理化学过程。
我们主要从器官系统和整体水平阐述家畜生理活动,有些内容稍涉及一些细胞水平的研究。
动物生理学的一些重要分支及其相关学科。
一)人体及哺乳动物的形态学(Morphology)和生理学
结构和功能在动物体内是不可分割的统一体,形态学研究与生理学研究保持密切联系。二者也是生物科学中最早发展的学科之一,19世纪中期,(法)ClaudeBernard奠定了人体生理学基础,当时生理学概念较为广泛,包括生理学和形态学,早期的形态学相当于现代所说的解剖学。由于研究的不断发展,解剖学与生理学分成了两个学科。以后由于组织学和细胞学等新型形态学科的发展,生理学研究也相应地向组织和细胞水平,但二者仍然保持密切关系,甚至使细胞学和细胞生理学重新融合成为细胞生物学。
二)比较生理学,生态生理学和进化Comparativephysiology, ecological physiology and evolution
Comparativephysiology 是揭示和比较异种动物生理活动的相同点和异同关,这种研究有助于阐明动物物种之间的亲缘关系,以及它们的进化史。
ecological physiology研究动物个体与环境之间的关系。
三)细胞生理学和普通生理学
Cellular physiology 研究细胞的结构和机能
General physiology 研究生物基本生理活动的原理和规律。
两个分支在许多方面相同,但普通生理学的研究领域比较广泛。生物的基本生理活动虽然主要在细胞内进行的,但有些超微细胞结构和细胞外的活动却只能包括在普通生理学范畴内。
四)生化和生物物理学biochemistry and biophysics
Biochemistry是研究生命物质的性质,反应和相互作用的科学,即揭示C内能量转化的代谢途径。
生物物理是研究生物体内各种生物物理过程的科学
例如:分析和检测动物的能理传递和转化,生物中产生和传播机理,物质运动的动力学规律。
二、目的和任务
家畜生理学是一门基础理化学科,先行课……后续课。
兽医专业:生理,病理解剖学、药理学,诊断学,内科学、产科学等。
畜牧专业:饲养和营养学、环境卫生学、繁殖学等。
这门学科是随着畜牧生产和兽医临床的需要而发展起来的,它的发展又为畜牧生产和兽医临床服务的。
通过学习生理学,不仅为后续课打基础,而且掌握正常的生命活动规律,指导畜牧生产。
如:瘤胃内,微生物可以合成B族V,配合饲料时应考虑—仔猪出生后,胃液缺乏HCI,一般在生后20天前后,胃液中才有少量HCI,胃内仅有凝乳酸,胃蛋白酶也很少,不能消化植物蛋白质。因为HCI可使蛋白质膨胀变性,同时提供酸性环境。使蛋白质易于消化,因此仔猪可以吃乳而不能过早地分解和利用植物性饲料。
在兽医临床上,掌握了正常的,才能理解和知道异常的。
如:正常血压、心率、血液的一些正常值,诊断时尿的生成:涉及滤过、重吸收、人泌和排泄、
药理:利尿药、双氢尿噻、抑制Na C1的重吸收从而增加水的排出(重吸收外)
病理情况下:发病时,少尿(急性肾小球肾炎,内皮C肿胀,基底膜增厚,滤过膜通透性减小,这涉及滤过问题)。
神经内分泌学—从动物生理学、动物化学发展成一门独立学科,近10年来发展迅速。
经典的动物生理学,主要研究动物机体的生命活动规律,,重点阐明神经及内分泌对各种生理过程的调节,而动物生理学的研究对象不但阐明有关动物的上述生理特点,并进一步如何调节控制有关生理过程以提高禽畜的生产力。
四. 生理学知识在动物生产中的应用
1.β—肾上腺素能受体激动剂—提高瘦肉率。
刺激β-受体促进体内脂肪分解神经潴留应用β-受体激动剂提高肉用畜禽瘦肉率即通过生理调控改善胴体的组成。
动物基因工程:
分两类:一、目的基因导入微生物或其它单细胞-蛋白质类物质,目的是提高畜牧产量,如生长激素,防治畜禽疾病的工程疫苗
二、基因导入动物的受精卵,培育出生产性能明显提高的转基因畜禽并能传给下一代。
1982美国大鼠的生长激素基因导入给小鼠-碩鼠。转基因猪早期死胎畸型。
例子:GH的N激素调控:
用基因工程生产出GH,牛、产奶量提高15—24%饲料利用率提高5—20%
猪、日增重提高12.6%饲料转换率17.5%体脂减少24.7% 瘦肉增加14.7%
问题:①严格的种间物异性
②只能口服,不便
③生产成本高
3.食欲促进剂:提高摄食量,促进生产
苯二氮化合物,乙磺氟安定可影响摄食中枢,摄食增3% 增重4% 料转化率8%
三、动物生理学的研究方法
动物生理学是一门实验科学,动物生理学的知识必须来自对生命现象的客观观察和实验。所谓观察,就是如实地把自然的客观现象记录下来,加以概括和统计,做出结论。例如,我们欲知健康成年牛的心搏频率,可以在一大群健康成年牛中进行观察,计数每头牛在安静状态下每分钟心搏次数。积累数千头牛的观察记录,加以统计分折,得出平均值为每分钟心搏若干次,于是得出结论:健康成年牛安静状态下心搏频率大致平均每分钟若干次。
所谓实验,就是人为地创造一定条件,使平时不能观察到的某种隐蔽的或微细的生理变化能够被观察,或某种生理变化的因果关系能够被认识。绝大多数生理学问题都要借助于这类方法进行研究。生理学所进行的动物实验方法,归纳起来不外乎慢性实验和急性实验两大类,而急性实验又可分为在体(in vivo)和离体(in vitro)两种。慢性动物实验方法主要是在无菌条件下对健康动物进行手术,暴露要研究的器官(如消化道各种造瘘手术)或摘除、破坏某一器官(如切除某一内分泌腺),然后在接近正常生活的条件下,观察所暴露器官的某些功能,观察摘除或破坏某器官后所产生的功能紊乱等。这种方法由于以完整动物为实验对象,又是在接近正常生活的条件下进行的,因此获得的结果比较接近正常生理活动规律。急性在体实验方法,是在无痛条件下剖开动物身体,对某一两个器官进行实验观察。这种方法比慢性实验方法简单,易于控制条件,有利于观察器官间的具体关系和分析某一器官功能活动的过程与特点,但与正常情况下的功能活动有一定差别。离体实验方法是在动物体内取出某一器官,置于人工的环境中,研究它的机能和有关因素的作用。例如,观察离体肌肉的收缩或离体心脏的活动等。但所得结果不一定能代表它在正常机体内的情况。
近二三十来,由于基础科学和新技术的迅速发展,以及相关学科间的交叉渗透,使生理学的研究得到迅速发展。有关器官机能的知识,特别是内分泌和中枢神经系统的机能的知识日新月异。如微量化学分析、免疫学、组织培养、电子显微镜、放射性同位素、PCR、电生理和电子计算机技术的应用,促使生理学从器官系统机能活动的描述,深入到细胞分子水平的研究。无创伤探测方法的应用,促进了整体水平的研究。
总之,所有这些不同的实验方法,各有其特殊的意义。在进行生理学研究时,应根据其研究的任务和课题的性质,选择最适当的方法。这里必须着重指出,无论采取哪种实验方法,在解释研究结果时,都必须持实事求是的态度,既不能把局限于某种特定条件下所获得的资料引伸为普遍性规律;更不能把一种动物实验的结果,不加区别的移用于所有的动物。这样才能正确、客观地反映事物的本质