周皮上的通气结构是皮孔
当细胞的伸长生长停止后,细胞壁仍可以继续生长加厚,这时生成的壁称为次生壁
由成熟组织细胞,经历生理上和形态上的变化,脱离原来的成熟状态(即脱分化),重新恢复细胞分裂能力而转变成的分生组织。木栓形成层是典型的次生分生组织
韭菜蒜、葱、的叶子剪去上部还能继续伸长,是因为叶基部居间分生组织活动的结果,
在水稻、小麦等谷类作物茎的节间基部保留有居间分生组织,其活动的结果,使茎节急剧伸长,以完成拔节和抽穗
细胞壁的构建受到的引导微管
种子,裸子植物和被子植物特有的繁殖体,它由胚珠经过传粉受精形成。种子一般由种皮、胚和胚乳3部分组成,有的植物成熟的种子只有种皮和胚两部分
细胞壁是植物细胞中最大的钙库。
果实成熟时果肉变软是由于果胶质被分解
质体可分为白色体、叶绿体和色质体
叶原基和芽原基在顶端分生组织的表面发生,这种起源方式成为外起源 维管射线是由射线原始细胞分裂,分化而成,因此,是次生结构,所以也称次生射线
禾本科植物种子均有胚乳
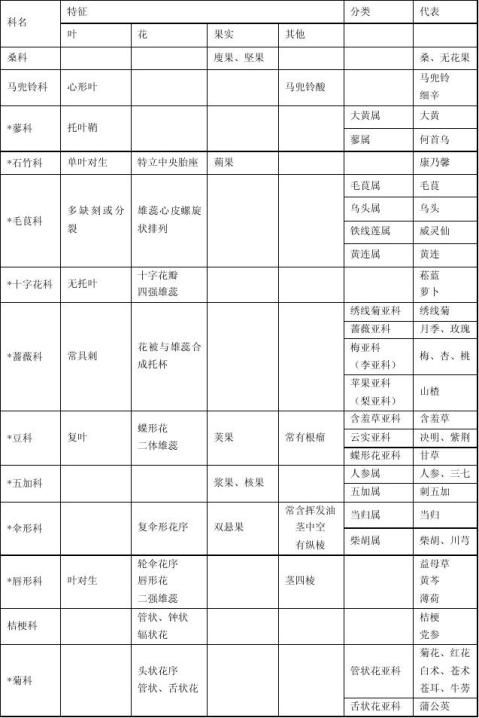
错.倒生胚珠发育的种子不具有种脊,直生胚珠发育的种子具有种脊
内表皮细胞排列整齐而紧密,在细胞的径向上、下壁和壁上,常有木质化和栓质化的加厚,呈带状环绕细胞一周,称凯氏带。
均为侧生分生组织,束中形成层具有初生分生组织的性质,束间形成层却是典型的次生分生组织
黄瓜、葡萄的卷须和豌豆的卷须依次是茎卷须,叶卷须
泡状细胞与叶内卷和折叠有关
眼子菜木质部退化,无机械组织的叶
根吸收的矿物质主要蒸腾作用靠向上输送
在花的组成中花托必不可少
柱状最原始的花托形状
柔荑花序毛白杨单性花、成熟后整个花序一起脱落
纤维层与花药开裂有关的结构是
花药壁完全分化时,自外向内的结构表皮,纤维层,中层,绒毡层
花药系统发育中,单核花粉的形成过程是孢原细胞——造孢细胞——花粉母细胞——小孢子
绒毡层的发育与植物的雄性不育有很大关系
双受精后初生胚乳核不经过休眠开始分裂,而合子需经过一段时间的休眠期才开始细胞分裂
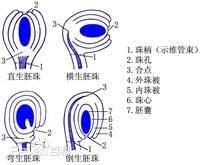
外胚乳来源于珠心
被子植物生活史中,配子体阶段始于大、小孢子 原核生物DNA没有组蛋白,真核生物有
裸子植物的小孢子叶又可称雄蕊

习见的植物的配子体称原叶体,虽能独立生活,但生活期短,跟孢子体相比,不占优势地位。种子植物的配子体即花粉粒和胚囊,(配子体所对应的雌雄配子分别为花粉粒——精子(雄配子),胚囊——卵细胞(雌配子)其中有关花粉粒致死基因典型代表为女娄菜)仅由很少细胞组成,不能独立生活,寄生在孢子体上。
胚乳来自雌配子体,是配子体世代
松属植物中叶是3针1束的是白皮松
松杉纲的植物茎的髓部小,次生木质部发达,由管胞组成,无导管,具有树脂道
买麻藤纲,次生木质部有导管,无树脂道。雌配子体无颈卵器,这些都是裸子植物中最进化类群的性状
典型被子植物的花由花托、花被 、花柄雌蕊群和雄蕊群组成 一朵花的花瓣若大小一致,形状也相同,而自花的中央向外呈辐射式排列,就称为「整齐花」也称「辐射对称花」 多为直根系和花粉常具有单个萌发孔分别是木兰纲和百合纲的性状
木瓜 不属于葫芦科
萝卜和胡萝卜分别属于十字花.伞形科 被子植物第一大科菊科,之后排兰科、豆科
向日葵头状花序的边缘花为舌状花
子房呈180度扭转的是兰科
木兰科托叶环
共质体 通过胞间连丝结合在一起的原生质体。

,输导有机物的为筛管和伴胞 次生结构 由维管形成层活动产生的区别于顶端分生组织形成的初生结构的结构
核相交替 生活史中单倍体核相和二倍体核相交替出现的现象。
二叉分枝 顶端分生组织平分为2个,各形成一个分枝,在一定时候,以这种方式重复分枝

部分构成的结构 单体雄蕊 雄蕊群花丝结合成一体,雄蕊多数花丝联合成管。
双受精 被子植物花粉粒中的一对精子分别与卵和中央细胞极核结合。 真果 仅由子房发育而形成的果实
成螺旋状排列在球茎的轴上的组成,雌球花的变态大孢子叶
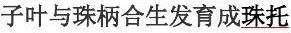
银杏的大孢子叶变成珠领有些红豆杉纲植物的大孢杨柳科

,花单性,无花被,具有由花被退化而来的花

具有真正的花 开花过程是被子植物的一个显著特征。被子植物的花由花被 (花萼、花冠)、雄蕊群和雌蕊群等部分组成。花被的出现,一方面加强了保护作用,另一方面增强了传粉效率,以达到异花传粉的目的
无限花序是指在开花期内,可随花序轴的生长,不断产生花芽,或重复地产生侧枝,每一侧枝顶上分化出花。这类花序的花一般由花序轴下面先开,渐次向上,同时花序轴不断增长,或者花由边缘先开,逐渐趋向中心。总状花序、圆锥花序、穗状花序、伞形花序等都属于这一类
有限花序为花序主轴顶端先开一花,因此主轴的生长受到限制,而由侧轴继续生长,但侧轴上也是顶花先开放,故其开花的顺序为由上而下或由内向外。
葇荑花序
一般整个花序一起脱落。
第二篇:苏州大学---药用植物学,被子植物知识点总结