一, 绪论-细胞概述
1,细胞学说(Cell Theory),所有的生物都是由一个或多个细胞组成的,细胞是生命的基本单位,一切细胞产自细胞。
2,原生质理论,有机体的组织单位是一小团protoplasm, 这种物质在一般有机体中是相似的
并把细胞明确地定义为:“细胞是具有细胞核(Nuclei)和细胞膜(Plasma membrane)的活物质”
3,细胞结构共性
选择透性的膜,遗传物质,核糖体
4,细胞功能共性
自我增殖和遗传物质复制,新陈代谢与酶学反应,运动性,应答与自我调节,复杂而有序,进化。
5,结构域 (domain)∶是指生物大分子中具有特异结构和独立功能的区域。
6,
7,
8,1
9,1
10, 1
11,
二, 细胞生物学研究方法
1,
2, 镜像形成的三要素:照明系统,样品,透镜系统 显微镜
◆相差显微镜(Ph2,标本密度不同导致光程差 振幅差
无色、透明、活细胞结构
◆微分干涉显微镜
更强立体感 ,便于显微操作
◆暗视野显微镜(丁达尔现象:光斜射到样品上)
物体轮廓,无微细结构
◆荧光显微镜(紫外光源激发细胞内荧光标记染料) ◆激光共聚焦显微镜
激光扫描、分辨力提高、可立体三维重建
电子显微镜
◆透射电子显微镜:细胞内部结构观察
◆扫描电子显微镜:样品表面形貌特征观察
◆扫描隧道显微镜
微观世界物质表面特征,但样品必须是导体
◆原子力显微镜
微观世界物质表面特征,不要求导体,提供三维表面图,不需样品处理、常压及液体环境中工作;但分辨率低于扫描电镜 3, 细胞组分的分析方法
细胞化学技术,细胞分选,细胞组分的分离
4, 细胞化学技术
◆酶细胞化学技术(酶与底物)
酶细胞化学技术就是通过酶的特异细胞化学反应来显示酶在细胞内的定位。
◆免疫细胞化学技术(抗原抗体)
利用免疫反应定位组织或细胞中的抗原成分分布 ◆放射自显影技术
利用放射性同位素标记研究细胞内大分子的合成动态(分布、定位、排出、合成等)
◆其他:DNA、RNA染色
原位杂交技术
5, 细胞分选
◆流式细胞术
干涉检测器:发现细胞
荧光检测器:发现标记
◆染色体分选
6, 细胞组分的分离
超速离心技术:分离细胞器与生物大分子及复合物
◆ 速度离心: 差速离心、移动区带离心
◆等密度离心:层析分离技术
层析分离技术:分离蛋白质
◆ 凝胶过滤
◆亲和层析
◆离子交换层析
7,细胞工程技术
细胞培养
细胞融合与单克隆抗体
基因转染与敲除
转染 (transfection):将具有生物功能的核酸转移或运送到细胞内并使核酸在细胞内维持其生物功能
8,组学研究技术
三, 细胞质膜与跨膜运输
1, Common functions of plasma membrane
界膜和区室化;功能定位与组织化。参与细胞间的相互作用;信号检测与传递。能量转换;调节运输
红细胞血影
是将分离的红细胞放入低渗溶液中,水渗入到红细胞内部,红细胞膨胀、破裂,从而释放出血红蛋白,所得到的红细胞质膜具有很大的变形性、柔韧性和可塑性,当红细胞的内容物渗漏之后、质膜可以重新封闭起来称为红细胞血影。
膜脂:甘油磷酯,鞘脂,胆固醇
膜脂的特性:膜脂的基本性质是“amphipathic molecule”, 能够self assembling as 双层结构或自我封闭成球状,受到损伤时可以自动再封闭。 2, 3,
脂质体(Liposome)
可用作生物膜的研究模型,并可作为生物大分子与药物的运载体。
膜脂的功能
◆构成膜的基本骨架,去除膜脂,则使膜解体;
◆是膜蛋白的溶剂,一些蛋白通过疏水端同膜脂作用,使蛋白镶嵌在膜上,得以执行特殊的功能;
◆膜脂为某些膜蛋白(酶)维持构象、表现活性提供环境, 一般膜脂本身不参与反应(细菌的膜脂参与反应);
◆膜上有很多酶的活性依赖于膜脂的存在。有些膜蛋白只有在特异的磷脂头部基团存在时才有功能。
4,膜糖
◆存在方式:glycoprotein (93%);glycolipid (7%)
◆膜糖功能
? 保护作用
? 细胞与环境的相互作用
糖脂
糖脂普遍存在于原核和真核细胞膜上,含量约占膜脂的5%
以下;
最简单的糖脂是半乳糖脑苷脂,它仅有一个半乳糖作为
极性头部;
变化最多、最复杂的是神经节苷脂,它是神经原质膜具特
征性的成分;
保护细胞膜、信号识别。
膜蛋白
◆存在方式:
整合蛋白:跨膜蛋白,部分或全部镶嵌在膜中
外周蛋白:外露在膜的外侧或内侧,非共价键
脂锚定蛋白:共价键方式与脂分子连接
◆膜蛋白功能
运输,代谢,连接,信号转导
去垢剂可以破坏脂双层溶解膜蛋白
利用温和去垢剂溶解、纯化和重建功能膜蛋白的系统
5, 膜的分子结构及特点
◆流动镶嵌模型 (Fluid mosaic model)
膜上脂质分子排成双层构成生物膜的骨架,蛋白质分子以不同方式镶嵌或联结于脂双层上;膜的两侧结构是不对称的,膜脂和膜蛋白具有一定的流动性。
◆脂筏模型(Lipid rafts model)
膜上一些特殊的脂质能形成稍厚的微区,这种大小约70纳米的动态结构像筏子一样,带着大量的膜蛋白以集中完成各种复杂功能,这就是生物膜的脂筏模型。
脂筏模型是对流动镶嵌模型的补充!!!!!
脂筏特点
具有较长的饱和脂肪酸链,分子间的作用力较强,所以这些区域结构致密,介于无序液体与液晶之间,称为有序液体(Liquid-ordered)。在低温下这些区域能抵抗非离子去垢剂的抽提,所以又称为抗去垢剂膜(detergent-resistant membranes,DRMs)。
像一个蛋白质停泊的平台,与膜的信号转导、蛋白质分选均有密切的关系。
脂筏中的胆固醇就像胶水一样,它对具有饱和脂肪酸链的鞘磷脂亲和力很高,而对不饱和脂肪酸链的亲和力低。用甲基-β-环糊精(methyl-β-cyclodextrin)去除胆固醇,抗去垢剂的蛋白就变得易于提取。
膜中的鞘磷脂主要位于外层,而且大部分都参与形成脂筏。 脂筏的面积可能占膜表面积的一半以上。脂筏的大小是可以调节的。小的独立脂筏可能在保持信号蛋白呈关闭状态方面具有重要作用;当必要时,这些小的脂筏聚集成大一个大的平台,在那里信号分子(如受体)将和它们的配件相遇,启动信号传递途径。 膜的不对称性(Membrane asymmetry)
研究方法:冰冻蚀刻技术 4,
?膜不对称性的表现:
膜脂不对称性:脂双层两小叶(leaflet)中分布的各类脂的含量不同。
膜蛋白的分布不对称:在膜两侧分布不对称,一般说,细胞质面的蛋白比外表面少,一些酶和受体多处于外表面。
膜糖的不对称:糖脂分布在动物细胞外表面,糖蛋白分布于脂膜双层的外叶。
?膜的不对称性的意义 膜脂、膜蛋白及膜糖分布的不对称性导致了膜功能的不对称性和方向性,保证了生命活动的高度有序性。
膜不仅内外两侧的功能不同, 不同区域的功能也不相同。造成这种功能上的差异,主要是膜蛋白、膜脂和膜糖分布不对称引起的。
细胞间的识别、运动、物质运输、信号传递等都具有方向性。这些方向性的维持就靠分布不对称的膜蛋白、膜脂和膜糖来提供。 膜的流动性(MEMBRANE FLUIDITY)
鼠、人细胞融合实验
?膜脂的运动方式
◆侧向扩散(lateral diffusion)
◆旋转(rotation)
◆翻转(flip-flop)
◆屈曲(flexion)
?影响膜流动性的因素:膜的流动性依赖于膜的组分和温度。
◆相变温度(transition temperature)
在生理条件下, 膜脂多呈拟液态(liquid-like state)。温度下降至某点, 则变为晶态(frozen crystalline gel)。一定温度下, 晶态又可熔解再变成液晶态,这种临界温度称为相变温度。
◆相变(phase transition)
在不同温度下发生的膜脂状态的改变称为相变。不同的膜脂由于成分不同而各有其相变温度。
◆膜脂肪酸链对膜流动性的影响:不饱和程度和链的长短 ◆胆固醇使得脂质在双层中堆积更为紧密,提高稳定性及有序性
可塑分子:
在相变温度以上,它可使磷脂分子的脂酰链末端的运动减小,即限制膜的流动性。
在相变温度以下,可增加脂类分子脂酰链的运动,这样可以增强膜的流动性。
◆卵磷脂/鞘磷脂比值对膜脂流动性的影响
卵磷脂所含的脂肪酸链的不饱和程度高,链较短,相变温度低,因此卵磷脂含量高,流动性大。
而鞘磷脂的脂肪酸链的饱和程度高,相变温度也高,因此,鞘磷脂的含量高,流动性低。
◆膜蛋白对膜脂流动性的影响
?膜的流动性的意义:
◆ 酶活性
◆ 物质运输
◆ 能量转换
◆ 信号转导
◆ 细胞周期
在M期, 膜的流动性最大, 而在G1期和S期, 膜流动性最低;
6, 物质的跨膜运输
脂双层具有半通透性质:
极性分子,只能很慢地穿过合成脂双层, 但膜上有一些特别的蛋白帮助这些分子跨膜运输。膜蛋白的主要功能就是跨膜运输营养分子、排出代谢废物以及调节胞内的离子浓度,这些跨膜运输蛋白占膜蛋白的15-30%。
?两类主要的跨膜运输蛋白
转运蛋白(transporter)和通道蛋白(channel)
被动运输(passive transport)
主动运输(active transport)
偶联运输:又称协同运输。这种运输需要先建立离子学梯度(Ion gradients),在动物细胞主要是靠Na+泵、在植物细胞则是由H+泵完成的。
ATP驱动的主动运输:利用ATP水解成ADP和磷酸所释放出来的能量跨膜主动运输物质的蛋白叫做ATP驱动泵,也称为转运ATP酶,它在原核细胞和真核细胞中广泛存在。
P型泵 - 钠钾泵:也叫做钠钾ATP酶,它利用ATP水解的能量逆电化学梯度将钠离子运出细胞、将钾离子运进细胞。
F型泵 - ATP合成酶:通常也被称作ATP合成酶,它们利用质子的跨膜梯度驱动ATP合成。
V型ATP泵与F型ATP泵结构类似,它们通常是利用ATP水解的能量来驱动质子运输,将质子泵进细胞器
ABC转运蛋白是膜转运蛋白中的一个大家族,跨膜转运种类繁多的分子,包括无机离子、氨基酸、单糖、多糖、小肽甚至是蛋白质。ABC转运蛋白有两个高度保守的ATP结合区(ATP-binding cassette),结合ATP可以使该区域二聚化,水解ATP使得这个结合区解聚。
光能驱动的主动运输
一些细菌和古细菌能利用光能作为能量来源,驱动跨膜的离子运输。它们通常需要一个特别的可以吸收光子的生色团分子(如蛋白质-细菌视紫红质(bacteriorhodopsin)的视黄醛(retinal)生色团),
四, 细胞表面与细胞互作
细胞表面
结构:细胞被(cell coat),细胞质膜(cytoplasmic membrane) 功能:保护细胞,使细胞有一个相对稳定的内环境;
参与细胞内外的物质交换和能量交换;
参与信号的识别和信息的传递;
参与细胞运动;
维护细胞的各种形态;与免疫、癌变等有十分密切关系。
?细胞被(cell coat)or多糖包被或糖萼(Glycocalyx)
组成:“糖”。功能:机械保护,信息交流
2, 细胞壁
?细胞壁的结构组成
(植物)细胞壁(多糖) = (动物)细胞外基质(蛋白质分子)
纤维素(cellulos): 葡萄糖多聚体
半纤维素(Hemicellulose):不同类型单糖的异质多聚体 果胶(pectin):半乳糖醛酸及其衍生物多聚体
木质素(lignin):聚合芳香醇
糖蛋白(glycoprotein):extensin
?植物细胞壁的分泌合成
◆中间层形成:主要是果胶。果胶是相邻两细胞壁所共有,并且
具有将两个细胞连接在一起的作用。
◆初生壁形成:是细胞正在生长时形成的。初生壁由纤维素、半
纤维素、果胶和糖蛋白等松散组成。
◆次生壁形成:是在细胞停止生长后分泌形成的。主要是增加细
胞壁的厚度和强度。次生壁主要成分是纤维素和
木质素。
?植物细胞壁的作用
提供机械强度
植物细胞壁是一个动态结构,能够进行很多活动:
3, 细胞外基质Extracellular matrix
细胞分泌的大分子在细胞间交织连接成网状结构,称ECM
维持细胞形态及活性,信号转导,细胞分化
?细胞外基质的组成∶蛋白聚糖,结构蛋白,黏着蛋白
蛋白聚糖(Proteoglycan,PG)
构成ECM的基质 1,
糖胺聚糖 (Glycosaminoglycans)
重复二糖单位构成无分支长链多糖
二糖单位之一是氨基己糖(氨基葡萄糖或氨基半乳糖), 故
称为糖胺聚糖。
二糖单位包括∶硫酸软骨素,透明质酸
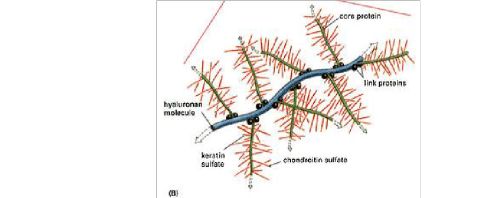
透明质酸(hyaluronic acid)
不同于其他PG,在细胞外基质中也可游离存在。作为长轴,
将蛋白聚糖连接在一起。
结构蛋白
?胶原(Collagen):不溶于水的糖蛋白,ECM的支柱蛋白。
胶原的作用:●胶原是骨、腱和皮肤组织中的主要蛋白,在
这些组织的细胞外基质中,胶原起着细胞外基质骨架作用;●胶原具有促进细胞生长的作用;●维持并诱导细胞分化的作用。
皮肤过度松弛症:胶原酶的遗传缺陷,胶原纤维不能正确地装配
?弹性蛋白(Elastin)
是弹性纤维(elastic fibers)的主要成分。
◆功能:能够赋予细胞外基质弹性。
黏着蛋白(adhesive protein)
细胞外基质与细胞联系的桥梁,促进细胞同基质结合 ?纤连蛋白(Fibronectin,FN)
?层连蛋白(laminin,LN)
4, 细胞外基质与细胞的相互作用
?基膜(Basement membrane )
◆细胞外基质特异区:上皮和内皮的基底面
?整联蛋白(integrin)
细胞外基质受体
受体浓度高,与配体结合力低
胞外端同基质蛋白结合
胞内尾部与肌动蛋白结合
细胞外基质与细胞内骨架双向联络,即细胞黏着
信号传递
5, 细胞识别,黏着,连接
先?细胞的识别 (Cell Recognition)
细胞对同种或异种细胞、同源或异源细胞以及对自己和异己分子
的认识和鉴别。
后?细胞的黏着(Cell Adhesion)
在细胞识别的基础上, 同类细胞发生聚集形成细胞团或组织的过程。
6, 细胞识别
抗原-抗体的识别
酶与底物的识别
细胞间的识别
基本特性:选择性-特异性!!!
?细胞识别与黏着的分子基础
糖-识别
蛋白-黏着
?识别反应内吞细胞黏着信号反应
7, 细胞黏着
概念:在细胞识别的基础上, 同类细胞发生聚集形成细胞团或组织的过程。
引起细胞黏着的主要是膜蛋白-细胞黏着分子
共有六种不同的方式的细胞黏着:四种细胞间黏着,两种细胞与细
胞外基质的黏着
8, 细胞连接
概念:细胞表面的特化结构,或特化区域。两个细胞通过这种结构连接起来。黏着在前,连接在后。
三种类型:◆紧密连接 (tight junction):封闭
◆斑形成连接 (plaque-bearing junction):黏着
◆间隙连接 (gap junction):通讯
紧密连接(tight junction)
?存在部位和结构特点
◆又叫不通透连接(impermeable junction), 它不仅连接相邻的细胞, 而且封闭细胞间隙, 使大多数分子难以在细胞间通透。 ◆这种连接方式普遍存在于腔道上皮细胞靠近管腔端的相邻细胞膜间。
◆从结构上看, 通过连接蛋白形成焊接线,封闭相邻细胞间的空隙。
?紧密连接的功能
◆连接作用
◆防止物质双向渗漏,
◆维持细胞的极性,并限制了膜蛋白在脂分子层的流动 斑块连接(plaque-bearing junction)
又称锚定连接(anchoring junction);主要靠黏着蛋白、整联蛋白和细胞骨架体系将相邻两细胞或细胞与细胞外基质连接在一起。
斑块连接分为:
●黏着连接(adherens junctions)
细胞间的连接作用与细胞骨架系统的肌动蛋白相关称为黏着连接。 (黏着带+黏着斑)
●桥粒(desmosome)
细胞间的连接作用与细胞骨架系统的中间纤维相关称为桥粒。 (桥粒+半桥粒)
间隙连接(gap junction)
一种跨膜结构,每个连接子由6个相同或相似连接蛋白(connexin)
亚单位环绕中央形成孔径为1.5-2nm的水性通道,相邻两细胞分别用各自的连接子相互对接形成分子间的通道,允许分子量在1200道尔顿以下的分子通过。
间隙连接的主要功能:
●机械连接作用;
●电偶联(Electrical coupling):
在神经冲动信息传递过程中起重要作用
●代谢偶联(Metabolic coupling)
代谢偶联可允许小分子代谢物和信号分子通过间隙连接 胞间连丝(plasmodesmata)
胞间连丝的结构∶
●是相邻植物细胞壁上的一个狭窄细胞壁通道
●有管状的内质网通过。因此,通过胞间连丝,使得相邻细胞的细胞质膜、细胞质、内质网交融在一起
●胞间连丝直径约30-60nm,允许1000道尔顿以下的分子渗透及大分子的运输 。
Communication Junction
◆一种特殊的细胞连接, 位于特化的具有细胞间通讯作用的细胞
◆具机械的细胞连接作用
◆在细胞间形成电偶联或代谢偶联, 以此来传递信号 ◆通讯连接的方式:
●间隙连接(gap junction):动物
●胞间连丝(plasmodesma):植物
五, 细胞通讯
细胞间或细胞内通过高精度和高效的接收信息通讯机制→→并通过放大引起快速的细胞生理反应/或引起基因活动,→→从而发生一系列的细胞活动来协调各组织行动,使之成为统一生命整体对外界环境变化作出综合应答。
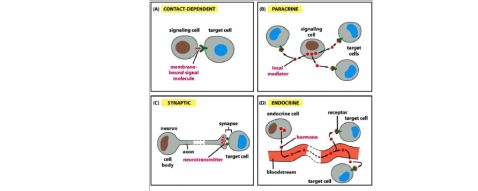
1, 信号分子
旁分泌信号,突触信号,内分泌信号,自分泌信号
2,
3, A,依赖接触b,旁分泌 c,神经元分泌 d,内分泌 受体 接收信息→受体,信号分子→配体(ligand) 细胞表面的受体
离子通道关联受体(受体本身就是离子通道的一部分)
也叫递质门控离子通道,受体是多次跨膜的蛋白质,受体与
配体结合后,构象改变,通道瞬间打开或关闭。
G-蛋白偶联受体(GTP结合的调节蛋白偶联)
酶关联受体(受体本身就是酶,又称催化受体)
多为一次跨膜的蛋白质,自身有酶的性质或者与酶结合在一起,受体外端具有配体结合部位,内端为催化部位,这类受
体大多为蛋白质激酶或者与蛋白质激酶结合在一起,他们被
激活后可使靶细胞中专一的一组蛋白质发生磷酸化。
胞内受体
存在于细胞质基质或核中,一些疏水性的信号小分子,如:
类固醇,甲状腺激素,类视黄素,维生素D,皮质醇等。
信号转导
细胞表面受体通过一定机制将外部信号转为内部信号。
包括:蛋白激酶,蛋白磷酸化酶
第一信使:细胞外部信号。
第二信使:由细胞表面受体转换而来的细胞内部信号。
第一信使同其膜受体结合后最早在胞膜内侧或胞浆中出现的4, 5,
仅在细胞内部起作用的信号分子;能启动或调解细胞内稍晚出现的细胞应答。
6, G-蛋白
组成:一般由三个亚基组成, 分别叫α、β、γ, β、γ两亚基通常紧
密结合在一起, 只有在蛋白变性时才分开。
功能位点:α亚基具有三个功能位点:①GTP结合位点; ②鸟苷三磷酸
水解酶(GTPase)活性; ③ADP-核糖化位点。
cAMP途径的信号解除和抑制
?信号解除
●通过磷酸二酯酶将cAMP降解,形成5'-AMP;
?信号抑制
●通过抑制型的信号作用于Ri, 然后通过Gi起作用。
?毒素对cAMP信号途径的影响
◆霍乱毒素(cholera toxin)
能把NAD+上的NAD-核糖转移到Gs蛋白的α亚基上,使G
蛋白核糖化(ADP-ribosylation),这样将抑制α亚基
GTPase活性??。
◆百日咳毒素(whooping couch toxin)
作用机理同霍乱毒素相同,但是使Gi蛋白进行ADP核糖化,
阻止Gi α亚基活化,其结果也是使cAMP的浓度增加。
磷脂肌醇信号途径
G蛋白偶联受体系统的一种。该通
路也称IP3、DAG、Ca2+信号通路,或称为PKC(Protein
kinase C)系统。
?Ca2+的第二信使作用
钙调蛋白(calmodulin)是Ca2+受体蛋白,结合后别构激
活,但并无酶活性。
? (PKC) IP3 /DAG/Ca2+信号的终止
DAG信号的解除
DAG只是由PIP2水解得到的暂时性产物, 寿命只有几秒钟,
靠两种方式进行降解:
◆被DAG磷酸激酶磷酸化,生成磷脂酸(PA),PA被转化为
CMP-磷脂酸,再与肌醇作用合成磷脂肌醇(PI)。
◆DAG 被DAG 酯酶水解生成单脂酰甘油,进一步水解成
自由的多不饱和脂肪酸和花生四烯酸甘油。
IP3作用的终止
◆IP3的水解
在5’磷酸酶的作用下, 水解为I(1,4)P2, 并进
一步水解成肌醇。5’磷酸酶是一种膜结合的酶。
◆在胞浆的肌醇磷酸脂3-激酶的作用下IP3被磷酸化成
I(1,3,4,5)P4。
Ca2+信号解除
●IP4参与打开细胞质膜上的Ca2+通道, 使细胞质中的
Ca2+较为持久地增高。
●胞内Ca2+浓度持久地升高, 可激活Ca2+-ATP酶(质膜、
内质网膜的钙泵),从而降低胞质中的 Ca2+,使胞质中的
2+-7 Ca迅速恢复到基态水平(10M),并使活性CaM-酶复合
物解离,从而酶失去活性,细胞反应终止。
7, 酶关联受体
特点:受体本身是一种具有跨膜结构的酶蛋白,其胞外域与配体
结合而被激活,通过胞内侧激酶反应将胞外信号传至胞内。
不需要信号偶联蛋白(G-蛋白), 而是通过受体自身的酪氨酸
蛋白激酶的活性来完成信号跨膜转换;
该通路对信号的反应比较慢(通常要几小时),并且需要许多细
胞内的转换步骤;
通常与细胞分裂相关。
8, 信号集成
趋同(convergent ): 不同的生长因子作用于不同的受体,
但能整合激活一个共同的效应物,如Ras或MAP激酶;
趋异(divergent): 相同配体,如EGF或胰岛素能够转换
激活许多不同的效应物,引起细胞的不同反应;
串话(crosstalk):不同信号途径间的相互影响
六, 细胞质,核糖体,RNAs
七,
八, 线粒体与叶绿体
线粒体
1,结构
?外膜
◆ protein : lipid =1:1
◆标志酶:单胺氧化酶 (monoamine oxidase);
◆外膜含有通道蛋白:孔蛋白 (porin);
?内膜
◆protein: lipid = 3:1
◆大量的心磷脂
◆线粒体进行电子传递和氧化磷酸化(oxidative phosphorylation)
的部位;
◆3 types of enzymes: transport, synthesis, electron transfer and ATP
synthesis
◆标志酶:细胞色素氧化酶
?膜间隙
◆标志酶:腺苷酸激酶(adenylate kinase)
◆功能: 核苷磷酸化: AMP+ATP?2ADP
?线粒体基质 (matrix)
◆标志酶:苹果酸脱氢酶
◆功能: ?
●代谢:丙酮酸氧化、TCA、beta氧化
●合成:DNA、RNA、蛋白质
?线粒体膜间间隙蛋白的定位
定位需要两个前导肽双导向序列
两种方式:保守性寻靶 ,非保守性寻靶
?线粒体内膜蛋白定位的三种途径
?外膜蛋白的定位
运送到外膜的蛋白质没有导肽, 它的氨基酸末端延伸部分就有识别受
体的功能,直接插入到外膜。
2,线粒体的功能:氧化磷酸化作用
?糖的有氧氧化(细胞氧化或生物氧化):
◆糖氧化成丙酮酸 (胞质)
◆丙酮酸脱羧生成乙酰CoA (线粒体)
◆乙酰CoA进入三羧酸循环彻底氧化 (线粒体)
◆葡萄糖酵解生成丙酮酸
细胞质中的葡萄糖(或糖原)在一系列酶的催化下生成丙酮酸的过程成
为糖酵解(glycolysis)。
●生物需要能量时首先利用多糖;
●必要时也会利用脂肪:
▲脂肪被水解生成脂肪酸;
▲脂肪酸能够进入线粒体基质,通过β氧化途径(β-oxidation pathway)循环氧化生成乙酰辅酶A。
3,呼吸链与电子传递
?线粒体能量转换策略
◆三羧酸循环中的能量转换
NAD+ → NADH FAD+ → FADH2
◆NADH和FADH2必须被氧化才能release energy
NADH + 1/2 O2 → NAD+ + 能量
FADH2 + 1/2 O2 → FAD+ + 能量
◆NADH和FADH2被氧化时释放的H+、电子和能量如何安置?
?呼吸链(respiratory chain)
◆又称电子传递链,是四种复合物组成的复合体
◆功能是参与对还原型辅酶的氧化
●H+的传递
●电子传递
?递电子体与递氢体
◆递电子体
◆递氢体
◆电子传递、氢质子传递方式
◆电子载体类型∶
铁硫蛋白、黄素蛋白、细胞色素和辅酶Q。
?主次电子传递链
◆主呼吸链∶
复合物Ⅰ、Ⅲ和Ⅳ构成主呼吸链,从NADH来的电子依次经过这三个复合
物, 进行传递。
◆次呼吸链∶
复合物Ⅱ、Ⅲ、Ⅳ构成次呼吸链,来自FADH2的电子不经过Ⅰ。
?F1-F0复合物是ATP合成部位
通过线粒体内膜重建实验证明位于内膜上的F1-F0颗粒是呼吸链中ATP
合成的部位。
? F0-F1颗粒的结构和功能
◆酶活性:两种酶活性
●ATP水解酶的活性
●ATP合成酶的活性
故F0-F1颗粒又称为ATP合酶(ATP synthase )。
?氧化磷酸化机制
化学渗透假说
1. 线粒体内膜上的呼吸链同时起质子泵的作用,可以在传递电子的同时将质子从线粒体基质腔转移到膜间腔2.线粒体内膜上的ATP合酶复合体也能可逆地跨线粒体内膜运送质子,一方面利用水解ATP的能量将质子从基质腔转移到膜间腔,另一方面当膜间腔存在大量质子使线粒体内膜内外存在足够的电化学H+梯度时,质子则从膜间腔通过ATP合成酶复合物上的质子通道进入基质,同时驱动ATP合成酶合成ATP;3.线粒体内膜本身具有离子不透过性,能隔绝包括H+、OH-在内的各种正负离子;4.线粒体内膜上有一系列可介导基本代谢物质和选择性转运无机离子进入线粒体内膜的载体蛋白。
叶绿体
1, 前质体(proplastid)∶直径约为1μm,或更小一些,它是由双层膜包被着未分化的基质 (stroma)所组成。
质体 (plastid):植物中的前质体随着在发育过程中所处的位置以及接受光的多少程度,分化成功能各异的质体(plastid),根据质体是否含有叶绿素和功能的不同,将质体分为白色体、叶绿体和有色体。
有色体(chromoplast)
不含叶绿素,含有叶黄素、胡萝卜素和类胡萝卜素,无类囊体结构。分布于高等植物的某些器官,如花瓣、果实和根细胞中,使其呈现黄色或桔黄色。不进行光合作用,功能是富集淀粉和脂类。
白色体(etioplast)
不含色素, 多存在于植物的分生组织和储藏组织中,具有创造和储藏淀粉、蛋白质和油脂的功能。
叶绿体 (chloroplast):
是植物细胞所特有的能量转换细胞器,其主要功能是进行光合作用,即利用光能同化二氧化碳和水,生成糖,同时产生分子氧。
含有叶绿素和胡萝卜素。有基粒、类囊体结构。分布于叶及茎皮层细胞中。 2, 叶绿体结构
叶绿体膜(chloroplast membrane)或称之为被膜(out envelope)、类囊体 (thylakoid)和基质(stroma)。
3种不同的膜:外膜、内膜和类囊体膜
3种彼此分隔的间隙:膜间间隙、叶绿体基质和类囊体腔。 3, 叶绿体蛋白的定位
?叶绿体蛋白的三种来源:
一是由叶绿体DNA编码,在叶绿体核糖体上合成
二是由核DNA编码,在叶绿体核糖体上合成
三是由核DNA编码,在细胞质核糖体上合成,通过导肽运输到叶绿体的不同部位
叶绿体基质蛋白定位∶两步运输:
第一步导肽与叶绿体的外膜受体结合;
第二步以转译后运输的机制(Post translation mechanism)跨过叶绿体的膜进入基质,导肽被金属蛋白酶水解掉。
4, 叶绿体功能 光合作用
光合作用反应式:
6CO2 + 6H2O+ 光 →C6H12O6 + 6O2 + 化学能 (674000千卡) 光合作用可分为两个不同的阶段:
即光反应和暗反应 (light and dark reaction)。
光反应是对光的吸收,将光能转变成化学能,并产生氧。 暗反应是对CO2的固定(从大气中摄取CO2并将CO2转变成含有还原碳的有机物)。
光反应
1、光能的吸收:原初反应
2、传递:电子传递
3、转换过程:光合磷酸化
光反应的场所是类囊体。
? 光系统 (photosystem)
进行光吸收的功能单位称为光合系统,由叶绿素、类胡萝卜素、脂和蛋白质所组成。
含有两个主要的成分∶
捕光复合物 (light -harvesting complex, LHC)
光反应中心复合物 (reaction-center complex)
固定一个光质子大约需要300个分子的叶绿素。
电子载体
光合作用的电子传递链(photosynthetic electron transfer
chain)是由一系列的电子载体构成的,同线粒体呼吸链中的电子载体的作用基本相似。
但二者不同的是,线粒体呼吸链中的载体位于内膜,而光合作用的电子载体位于类囊体膜并且是一个吸热的过程而不是放热的过程。
5, 光合磷酸化
在叶绿体进行的光合作用过程中, 会产生电化学氢质子梯度,因而能够驱动ADP磷酸化合成ATP,这一总过程由于是光驱动的,故叫光合磷酸化。
区别于线粒体的氧化磷酸化∶
氧化磷酸化是由高能食物分子氧化驱动,
光合磷酸化是由光质子驱动。
光合磷酸化的类型∶
非循环式光合磷酸化
循环式光合磷酸化
假循环式光合磷酸化
光合磷酸化的机理
化学渗透学说的基本要点是:在电子传递和ATP形成之间,起偶联作用的是膜内外之间存在的质子电化学梯度。电子在膜上的传递象一个H+泵,促使H+穿过膜进行定向传递,形成膜两侧的H+质子梯度。由于H+的传递是由泵来推动的,因此,在形成质子电动势梯度时就会储存能量,它可用于将ADP+Pi形成ATP。
九, 内膜系统与蛋白质分选
1,◆膜结合细胞器 (membrane-bounded organelles):
细胞质中所有具有膜结构的细胞器
◆细胞质膜系统
细胞内在生物发生上与质膜相关的细胞器:
质膜+内膜 ◆内膜系统
内膜系统是真核细胞特有的结构,主要包括内质网、高尔基复合体、溶酶体、各种转运小泡以及核膜和过氧化物酶体等功能结构
2,内膜系统的研究方法
放射自显影,GFP,亚细胞分离,突变
3,内质网 ER
4,RER与蛋白质合成
?信号序列的引导作用
◆与生俱来的三种信号序列
●寿命信号
●加工信号
●定位信号
▲核定位信号
▲引导肽
▲信号肽
?信号肽的特性
◆序列特征
15-35个氨基酸残基,N端1-多个正电荷AA,其后含有6-12个疏水残基
◆特异性:信号序列无特异性
◆位置:
●N-端,进入RER后被切除
(区别于内部信号肽)
?信号假说(signal hypothesis)
◆合成始于游离核糖体;
◆N端信号序列靠SRP/DP识别与内质网膜接触,然后插入内质网
的膜;
◆蛋白质以袢环形式穿过内质网的膜;
◆蛋白的存在方式:
●若信号序列被信号肽酶切除:完全进入腔
●若是停止转移信号: 膜蛋白
信号斑(signal patch)
是溶酶体酶形成的一个特殊的空间结构,信号斑是由几段信号肽形成的一个三维结构的表面, 这几段信号肽聚集在一起, 形成一个斑点被磷酸转移酶识别。
发动蛋白(dynamin)
发动蛋白在网格蛋白小泡形成过程中同出芽的颈部结合,一旦小泡装配完成,发动蛋白立即水解其本身结合的GTP从而将小泡与质膜切离。
十, 细胞骨架与细胞运动
1, 细胞骨架的组成和分布
◆微管主要分布在核周围,并呈放射状向胞质四周扩散;
◆肌动蛋白纤维主要分布在细胞质膜的内侧和细胞核膜的内侧; ◆中间纤维则分布在整个细胞中。
2, 细胞骨架的功能
◆作为支架(scaffold)
◆在细胞内形成一个框架(framework)结构
◆为细胞内的物质和细胞器的运输运动提供机械支持
◆为细胞的位置移动提供力
◆为信使RNA提供锚定位点,促进 mRNA 翻译成多肽
◆是细胞分裂的机器
◆参与信号转导
3, 微管
◆微管是由微管蛋白异源二聚体为基本构件,螺旋盘绕形成微管的壁
◆在每根微管中二聚体头尾相接, 形成细长的原纤维(protofilament)
◆13条原纤维纵向排列组成微管的壁
?微管蛋白(tubulin)
◆微管蛋白类型:
◆α和β微管蛋白形成长度为8nm的异源二聚体
◆每一个微管蛋白二聚体有两个GTP结合位点
● α亚基GTP结合位点
●β亚基GTP结合点是可交换位点(exchangeable site)。 ◆γ-微管蛋白的功能是帮助α、β微管的聚合。
?微管装配的动力学现象
◆踏车现象(treadmilling)
又称轮回现象, 是微管组装后处于动态平衡的一种现象。
◆动态不稳定性(dynamic instability)
微管随反应体系中游离α、β二聚体的浓度变化而发生的生长状态和缩短状态
的转变。
?影响微管稳定性的药物
◆紫杉醇(taxol)
存在于红豆杉属植物中的一种复杂的次生代谢产物, 促进微管聚合和稳定已聚
合微管的药物。
◆秋水仙素(colchicine)
秋水仙素与未聚合的微管蛋白二聚体结合, 阻止聚合。
?MAPs(微管结合蛋白)的功能
◆使微管相互交联形成束状结构;
◆促进微管的聚合;
◆作为分子发动机转运细胞物质的轨道;
◆提高微管的稳定性;
◆MAPs同微管的结合能够控制微管的长度,防止微管的解聚。
?微管的功能
支架作用
细胞内物质运输的轨道
◆触突运输(axonal transport)
◆鱼的色素细胞(fish pigment cells)
作为驱动蛋白与动力蛋白运输轨道
作为色素颗粒运输轨道
微管介导的物质运输
组成纤毛和鞭毛的轴丝
纤毛和鞭毛的运动机制:微管滑动模型
纺锤体和染色体运动
4,分子发动机的类型
●肌球蛋白(myosins)家族
●驱动蛋白(kinesins)家族
●动力蛋白(dyneins)家族
?分子发动机移动的主要特点
●驱动蛋白:从(-)端向(+)端的运输
●动力蛋白:从(+)端向(-)端运输
5, 动力蛋白
功能
●参与细胞分裂
●运输小泡和各种膜结合细胞器
6,微丝
又称肌动蛋白纤维(actin filament)
?微丝的形态结构
◆F-actin:
●F-肌动蛋白呈双股螺旋状,直径为8nm,螺距37nm。
◆G-actin:
三个结合位点:
●一个ATP结合位点
●两个肌动蛋白结合蛋白的结合位点。
?肌动蛋白纤维装配
◆ATP的作用
Actin = ATPase tubulin = GTPase
◆装配过程
●成核(nucleation)
●延伸(elongation)
●稳定状态(steady state)
?影响装配的因素
◆G-actin临界浓度
◆ATP的作用
◆离子的影响
2+++●在含有Ca, 以及很低的Na、K等阳离子的溶液中,
微丝趋向于解聚成G-肌动蛋白。
●在Mg2+和高浓度K+或Na+的诱导下, G-肌动蛋白则装配成纤维状肌动蛋白。
?微丝的动态性质
◆极性
◆踏车现象(treadmilling)
◆微丝的动态平衡
?作用于微丝的药物
◆细胞松弛素B(cytochalasins B)促进微丝解聚 ◆鬼笔环肽(phalloidin)抑制微丝解体
?微丝发机蛋白:肌球蛋白(myosin)
◆肌球蛋白的结构
由一个重链和几个轻链组成,并组成三个结构域∶
●头部: 与肌动蛋白、ATP结合的位点,负责产生力。 ●颈部: 同钙调素或类似钙调素的调节轻链亚基的结合来调节头部的活性。
●尾部:决定尾部是否同膜结合还是同其它的尾部结合的
位点
肌球蛋白运动的机理:滑动模型
?微丝在非肌细胞中作用
◆参与细胞的连接
◆细胞内运输作用
◆细胞质流动(cytoplasmic streaming)
◆细胞爬行(cell crawling)
7,肌收缩机制--滑动纤维模型
◆实验观察∶
在肌肉收缩过程中:
●肌节几乎缩短50%,
●A带的长度并没有发生变化
●I带几乎消失
◆机理:
滑动丝模型(sliding filament model)
?Ca2+在肌收缩中的作用
◆Ca2+释放
●运动神经信号引起肌细胞膜电位去极化
●T管(transverse tubules,T tubules) ∶
●运动神经信号通过T管打开Ca2+通道
◆Ca2+对肌收缩的调节作用
●Ca2+与肌钙蛋白结合解除原肌球蛋白的抑制作用
●肌动蛋白与肌球蛋白头部结合形成交联桥
●利用ATP进行滑动
8,中间丝 IFs
◆真核细胞中第三种细胞骨架成分
◆这种纤维的平均直径介于微管和微丝之间,故称为中间纤维 ◆它的直径为10nm, 故又称10nm 纤维
◆由长的、似杆状的蛋白装配而成
功能
◆给细胞提供机械强度
◆参与细胞连接
◆维持细胞核的形态
十一,细胞核与染色质
1, 核被膜
?功能
◆基因表达的时空隔离
◆核膜成为保护性屏障, 使核处于一微环境
◆染色体的定位和酶分子的支架
◆物质运输
?结构
◆外核膜 (Outer nuclear membrane):
内质网膜特化区
◆内核膜 (Inner nuclear membrane)
◆核周腔 (Perinuclear space)
两层核膜之间的空隙, 宽15-30nm,
其中充满无定形物质
◆核纤层 (lamina):核纤层蛋白 (lamin! Not laminin),支撑核被膜
◆核孔复合物 (nuclear pore complex)
◆由内外两层膜的局部融合而成
◆核孔的直径为80-120nm,沟通核质与胞质的开口。 ◆一个典型的哺乳动物的核膜上有3000-4000个核孔,相当于每平方微米的核膜上有10-60个。
◆合成功能旺盛的细胞,核孔的数量较多。
2, 核孔复合体的结构与功能
?核孔复合体的结构:鱼笼模型
◆胞质环
(cytoplasmic ring)
◆细胞质颗粒
(cytoplasmic granular)
◆核质环
(nucleoplasmic ring)
◆中央运输蛋白
(central transporter)
◆辐条 (spoke)
◆核被膜网格
(Nuclear Envelope lattice)
◆笼状体 (basket)
?核孔运输特点
◆被动运输
◆主动运输
●信号引导
●双向性
?核蛋白运输机制
◆核蛋白(nuclear protein):胞质中合成,但在核内起作用 ◆核定位信号(nuclear localization signals, NLS):一种信号序列,介导核蛋白入核。带正电的核心序列。 ◆核输出信号(nuclear export signals, NES):
◆输入蛋白(importin):水溶性的NLS受体
◆输出蛋白(exportin):识别NES
3, 染色质
◆染色质(chromatin)
◆染色体(chromosome)
◆染色体与染色质比较
●在化学本质上没有差异
●在构型上不同
●是遗传物质在细胞周期不同阶段的不同表现形式。 ◆人工染色体 (artificial chromosome)
●人工构建的含有稳定染色体的天然结构序列,即ARS、CEN、TEL序列的微小染色体,
●可以象天然染色体一样在寄主细胞中稳定复制和遗传,称为人工染色体。
?组蛋白的化学修饰
虽然组蛋白分子的氨基酸序列是高度保守的,也会进行一些化学
修饰,如酰基化、甲基化、磷酸化,以及ADP核糖化等。 ?核小体的结构特点
Nucleosome is basic structural unit of chromatin 。 ◆由200个左右碱基对的DNA和四种组蛋白结合而成;
◆其中四种组蛋白(H2A、H2B、H3、H4 )各2分子组成八聚体的小圆盘,是核小体的核心结构;
◆146个碱基对的DNA绕在小圆盘外面1 3/4圈。每一分子的
H1与DNA结合, 起稳定核小体结构的作用;
◆两相邻核小体之间以连接DNA(linker DNA)相连, 长度为~80bp不等。
4,染色体
◆核小体 (nucleosome)
◆螺线管 (solenoid)
◆超螺线管 (supersolenoid)
◆染色体 (chromosome)
从核小体开始到染色体, DNA总共压缩: 7倍6倍40倍5倍
DNA→核小体→螺线管→超螺线管→染色单体
灯刷染色体(lampbrush chromosome)
是卵母细胞进行减数第一次分裂时,停留在双线期的染色体。 它是一个二价体, 含4条染色单体。它 由轴和侧丝组成,形似灯刷。
5, 分子伴侣的概念及其特点
●由不相关类的蛋白质组成的一个家系
●它们介导其它蛋白质的正确装配
●但自己不成为最后功能结构中的组分。
◆该概念有以下特点:
●凡具有“介导”功能的蛋白,都称为分子伴侣,可以是完全不同的蛋白质;
●作用机理尚不清楚,故用“介导”二字,伸缩性较大; ●分子伴侣一定不是最终结构的组成部分,但不一定是一个分离的实体;
●装配的涵意比较广,包括:帮助新生肽的折叠,越膜定位, 亚基组装等。
?分子伴侣结构上的共同特点:
◆家族成员具有高度保守性
◆家族成员结构上具有相似性
◆大部分在体内为组成型表达,在刺激条件下会被进一步诱导。
◆被底物激活和增强的弱ATP酶活性
?分子伴侣的功能
◆帮助蛋白质折叠和装配
◆蛋白质的转运和定位
◆参与细胞器和细胞核结构的发生
◆应激反应
◆参与信号转导
十二,细胞周期
细胞周期是指连续分裂的细胞从一次有丝分裂结束后开始生长到下次有丝分裂终止所经历的全过程。在这一过程中,细胞的遗传物质进行复制并均等地分配给两个子细胞。
1, 细胞周期时相及类型
?Phases 阶段of the cell cycle
◆G1期 (Gap 1 phase),即从M期结束到S期开始前的一段间
歇期;合成前期,合成rRNA、蛋白质、脂、糖
◆S期,即DNA合成期 (DNA synthetic phase);DNA 合成期,组蛋
白,DNA复制所需的酶
◆G2期 (Gap 2 phase), 即DNA合成后 (S期) 到有丝分裂前
的一个间歇期;DNA合成后期,ATP,RNA、细胞分裂相关蛋白质
◆M期,即有丝分裂期 (mitosis phase)。不一定每种细胞都有四个时期,如胚胎细胞没有G1期。
G0细胞
又称休眠细胞。暂时脱离细胞周期,不进行增殖,也叫静止
细胞群,如某些免疫淋巴细胞, 肝,肾细胞等。
2, 细胞周期的研究方法
?细胞同步化 (synchronization)
◆选择同步法 (selection synchrony)
●有丝分裂选择法
●细胞沉降分离法
◆诱导同步法
●DNA合成阻断法:胸腺嘧啶阻断法
●中期阻断法:微管解聚药物
◆条件突变 (conditional mutants):酵母
3, 蛋白激酶在细胞周期调控中的作用
?细胞融合实验
◆研究者:19xx年,Colorado 大学的Potu
Rao 和 Robert Johnson
◆研究思路
◆研究方法
◆使用的细胞系
◆预测结果
G1期细胞与M期细胞融合
S期细胞与M期细胞融合
G2期细胞与M期细胞融合
成熟促进因子(maturation promoting factor,MPF),早期称为M-期促
进因子(M-phase promoting factor, MPF),是指M期细胞中存在的促进细胞分裂的因子
?细胞周期蛋白的鉴定
◆研究思路
◆实验设计(注意关键步骤的用意!!!)
●获得同步化的受精的海胆卵细胞
●在有放射性氨基酸的培养液中培养
●每10分钟取一次样分离纯化蛋白质进行分析
◆实验结果
●发现了周期蛋白B(cyclin B):如何发现?
●周期蛋白B的cDNA克隆与周期蛋白B的鉴定
?MPF的结构组成
◆是由两个不同的亚基组成的异质二聚体:
●催化亚基
▲是丝氨酸/苏氨酸型蛋白激酶
▲其活性有赖于周期蛋白,故蛋白称为周期依赖性蛋白激酶
(cyclin-dependent protein kinases, Cdks);
●调节亚基:周期蛋白(cyclin)
?有丝分裂的退出:周期蛋白B的降解
?遍在蛋白与周期蛋白的降解
?真核生物细胞周期调控的一般模型
◆三类周期蛋白-CDK复合物:
●G1期周期蛋白-CDK复合物
●S期周期蛋白-CDK复合物
●有丝分裂周期蛋白-CDK复合物
◆三个关键的过渡
●G1期→S期
●中期→后期
●后期→末期及胞质分裂期过渡
4, 哺乳动物细胞周期的控制
?哺乳动物细胞周期的限制点
?哺乳动物细胞周期受多种CDKs和周期蛋白的调控
◆细胞周期受一个小型Cdks家族的调节:
●CDK1、CDK2、CDK3、??等;
◆多种周期蛋白:
●周期蛋白A:S期和M期周期蛋白
●周期蛋白B:M期周期蛋白
●周期蛋白D1、D2、D3:G1中周期蛋白
●周期蛋白E:晚G1期、S期周期蛋白
5, 细胞周期监测点
与细胞分裂有关的基因称为细胞分裂周期 (cell division cycle, cdc) 基因。这些
基因表达的有序性, 受一些控制系统的监测:
?细胞周期中的三个主要关卡
◆G1关卡(靠近G1末期)(START或限制点):
◆G2关卡(在G2期结束点)
◆中期关卡(在中期末)
在每一个关卡,由细胞所处的状态和环境决定细胞能否通过此
关卡,进入下一阶段。
?MPF成熟促进因子的作用机制
◆促进染色体凝集
●H1组蛋白磷酸化
●H3组蛋白磷酸化
◆核纤层磷酸化
◆核被膜装配
◆细胞相关的酶与蛋白质磷酸化
MPF激活肌球蛋白促进胞质分裂
6, 有丝分裂
前期(Prophase)主要特征
四个主要事件:
染色体的凝集、
分裂极的确定、
核仁的消失、
核膜的解体。
前中期 (prometaphase)
纺锤体(spindle)的装配:
●纺锤体侵入细胞核的中心区,一部分纺锤体微管的自由端最
终结合到着丝点上,形成动粒微管。
●染色体剧烈地活动,个别染色体剧烈地旋转、振荡、徘徊于
两极之间。
中期
●该期主要特点是姐妹染色单体位于赤道板上,着丝粒分别被两端的中心体
发出的纤维连接
后期 (anaphase)
这一时期的主要特点是:着丝粒分开,染色单体移向两极。
末期
该期的主要特点是:染色体解螺旋形成细丝,出现核仁和核膜。
?纺锤体微管运动机理
微管去聚合作用假说
动粒微管不断解聚缩短,造成将染色体拉向两极。
纺锤体微管滑动假说
极-极分离是由极微管的两种不同类型的变化引起的。
7, 减数分裂
前期I的5个阶段
●主要事件是完成同源染色体的配对,此过程中要发生配对同源染色体间的分子重组
●该期细分为细线期、偶线期、粗线期、双线期、终变期
◆细线期 (leptotene stage,leptonema),又称凝集期 (condensation stage)
此期主要特点:
染色体已加倍,并凝缩成细线状,但看不到染色体的双重性。
◆偶线期 (zygotene stage, zygonema)
又称配对期 (Pairing stage),
此期的主要特点是:
●联会与联会复合体
▲联会 (synapsis)
▲联会复合体 (synatonemal complex)
●二价体与四分体
▲二价体(bivalent)
▲四分体 (tetrad)
◆粗线期 (pachytene stage, pachynema)
又称重组期 (recomination stage),
主要特点:
●同源染色体之间发生DNA片段的交换, 产生重组的基因
组合
●持续时间长
●可见重组节
◆双线期 (diplotene stage)
又称合成期 (synthesis stage),
主要特点:
●同源染色体分开,明显可见四分体;
●出现交叉
◆终变期 (diakinesis)
又称再凝集期 (recondensation stage)。
此期的主要特点:
●染色体变成紧密凝集状态;
●大多数核仁消失,交叉出现端化, 姐妹染色体借着着丝
粒连接在一起。
十三,胚胎发育与细胞分化
1, 配子发生与受精
◆配子发生(gamete genesis)
雌雄生殖细胞的发生都要经过三个时期, 即增殖期、生长期和成熟分裂期。
◆受精 (fertillization)
受精是成熟的精子与成熟的卵子相遇,两者融合重新形成一个2倍体细胞,即受精卵。
经过:
顶体反应(acrosomal reaction)、
皮层反应(cortical reaction)、
原核融合(pronuclei fussion)
等过程,受精作用结束。
2, 胚胎形成与分化
受精后,受精卵经过多次有丝分裂,形成许多分裂球,这个过程叫卵裂
◆ 细胞分化(differentiation)
细胞分化是指同一来源的细胞逐渐发生各自特有的形态结构,生理功能和生化特征的过程。细胞分化时的主要特征是细胞出现不同的形态结构和合成组织特异性蛋白质,演变成特定表型的细胞类型。
3,
4,
5, ◆分化细胞的特点 ? 个体中所有不同种类的细胞的遗传背景完全一样。 ? 分化细胞彼此之间在形态、结构、功能方面的不同是由于其拥有不同的蛋白质所致。 ? 细胞分化中最显著的特点是分化状态的稳定性。 ? 虽然细胞分化是一种相对稳定和持久的过程, 但是在一定的条件下, 细胞分化又是可逆的。 ◆ 细胞决定(cell determination) 细胞决定是指在细胞发生可识别的形态变化之前,就已受到约束而向特定方向分化。是分化潜能逐渐限制的过程,决定先于分化。 ◆ 参与细胞决定,影响细胞分化的因素: ?细胞质的作用 ?细胞的相互作用的影响 ?形态发生:(时间+空间) ,位置信息 ?其它:核质关系、细胞黏着、激素、环境如温度、营养、个体大小等 形态发生:(时间+空间) 胚胎发育不仅需要将分裂产生的细胞分化成具有不同功能的特异的细胞类型,同时要将一些细胞组成功能和形态不同的组织和器官,最后形成一个具有表性特征的个体,这一过程称形态发生 ◆转决定 (transdeterminination) 不按已决定的分化类型发育,而生长出不是相应成体的结构。它是一群细胞而不是单一细胞发生变化。 分化细胞的基因表达 ◆看家基因(House-keeping gene)∶是维持细胞最低限度功能所不可少的基因,如编码组蛋白基因、编码核糖体蛋白基因、线粒体蛋白基因、糖酵解酶的基因等。 ◆组织特异性(Tissue-specific gene)表达基因∶或称为奢侈基因(luxyry gene)。这类基因与各类细胞的特殊性有直接的关系,是在各种组织中进行不同的选择性表达的基因。 细胞分化主要是奢侈基因中某种(或某些)特定基因的选择性表达的结果。 基因表达调控 ◆ 基因组调控:DNA甲基化 ◆ 转录水平的调控∶顺式调控元件/反式作用因子 ◆转录后加工的调控: ◆ 翻译水平的调控: ◆ 翻译后加工 ◆ 其它:mRNA运输的控制、mRNA降解的控制等 果蝇发育的基因控制
三类基因控制着果蝇的发育:
◆母体效应基因(maternal-effect gene)
基因在果蝇卵母细胞成熟过程中由抚育细胞转录,合成的mRNA运送至卵母细胞,其翻译产物在早期胚胎发育中调节合子基因的转录。
这些基因决定果蝇的极性,即果蝇的头部、尾部、背部-腹部的轴。
卵细胞质母体信息( maternal information)
受精激活后,受精卵中蛋白质合成迅速增加,此时加入RNA合成抑制剂不能影响蛋白质合成,表明合成蛋白质的mRNA是卵母细胞带来的,并且是非活性状态存在,只有受精后才被激活,这些由卵母细胞带来的信息叫母体信息。
同源异形选择基因(homeotic selector genes)
在高等真核生物中,经常发生某一器官异位生长的现象,称为同形异位作用。同形异位作用是由一类叫同形异位基因的突变所产生。在目前已知的300多个同形异位基因中,都存在一个高度保守的同源区域,长180bp,编码60个氢基酸。这个同源域称为同源异形盒(homeobox),因此同形异位基因又叫做同源异形盒基因。
十四,细胞衰老、死亡与癌变
1, 衰老的概念
衰老(senescing,aging)是机体在退化时期生理功能下降和紊乱的综合表现,是不可逆的生命过程。细胞的衰老与死亡是新陈代谢的自然现象。
机体的衰老与细胞的衰老相关联。
现代人类面临着3种衰老:
◆生理性衰老
◆病理性衰老
◆心理性衰老
细胞的寿限
19xx年,Hayflick 首次报道了
体外培养的人成纤维细胞具有增殖分裂的极限,且分裂能力与个体的年龄有关。
来源于胚胎 → 分裂传代50次后开始死亡
来源于成年组织 → 培养15~30代就开始死亡
动物体细胞在体外可传代的次数,与物种的寿命有关。
? Hayflick界限 (Hayflick life span):
细胞至少是体外培养细胞的增殖能力不是无限的,而是有一定界限,细胞的衰老控制着细胞的分裂次数, 进而控制着细胞的数量。 ?细胞的寿限:
各类细胞本身的寿命差别显著, 一般说来, 能够保持持续分裂能力的细胞是不容易衰老的。分化程度高又不分裂的细胞寿命是有限的。 细胞衰老的特征
◆细胞内水分减少
◆色素生成和色素颗粒沉积
◆衰老过程中细胞质膜的变化:
流动性降低;兴奋性降低;配体受体复合物形成下降
◆衰老过程中线粒体的变化
数量减少,体积变大,膜破坏,DNA突变
◆细胞核的变化
核膜内折;染色质固缩;端粒缩短
◆细胞骨架的变化
◆蛋白质合成的变化
合成速度降低,蛋白定位改变
细胞衰老的理论
◆细胞衰老的线粒体损伤论 (Mitochondria ageing)
◆自由基理论:氧自由基 (Reactive oxygen species (ROS))
◆细胞衰老的端粒假说 (Replicative senescence and telomere shortening)
◆细胞衰老的表观调控: (Epigenetic regulation of senescence) ◆衰老的进化论
衰老的突变积聚、互逆多效理论
2, 细胞死亡
细胞死亡概念:
细胞死亡的一般定义是细胞生命现象不可逆的停止。细胞死亡有两种形式:一种为坏死性死亡,另一种为程序性死亡。
◆程序性细胞死亡的过程
●死亡激活期 (activation phase):接收

death signal
●死亡执行期 (execution phase):执行一套死亡程序
细胞凋亡的机理:基因调控作用的结果
●决定死亡的两个基因,即ces-1(ces表示CE细胞存活的调控基因)和ces-2
基因
●执行死亡的4个基因:ced-3、ced-4、ced-9和egl-1基因:“死亡机
器”(death machinery)
●7个与死亡细胞被吞噬细胞所吞噬的基因,即ced-1、ced-2、ced-5、ced-6、
ced-7、ced-10和ced-11。
●ced-9可抵消ced-3和ced-4的作用,防止细胞被杀死,因此是存活因子; ●死亡细胞在吞噬体中被降解的基因
?Caspase
◆自杀性蛋白水解酶是天冬氨酸特异性半胱氨酸蛋白酶(cysteine-containing
aspartate specific protease),简称caspase;
◆caspase-3、6、7和8 在FAS/TNF介导的程序性细胞死亡途径中起作用; ◆caspase-9和3 一起参与线粒体中Apaf-I、细胞色素c介导的程序性细胞死
亡;
◆在人类,已经鉴定了10种不同的caspase。
◆能够被caspase切割的靶蛋白
●蛋白激酶
●核纤层蛋白
●细胞结构蛋白
●与DNA修复相关的酶类
●caspase激活的DNase抑制蛋白
2,癌生物学
癌细胞的生物学特性:无限增殖
侵袭及转移
癌细胞恶性程度越高,分化程度越低
?癌基因与原癌基因
◆癌基因(oncogene):癌基因是细胞加速器,它们编码的蛋白使细胞生长不受
控制,并促进细胞癌变
●细胞癌基因 (c-onc):由细胞原癌基因突变而来;
●病毒癌基因 (v-onc):大约已经鉴定了100多种不同的癌基因,它们中的大
多数属于RNA肿瘤病毒基因 组中的基因。
◆原癌基因(proto-oncogene):
原本是细胞的正常基因,它们编码的蛋白质在正常细胞中通常参与细胞的生长
与增殖的调节。但突变后成为促癌的癌基因
(cancer-promoting oncogene),导致细胞癌变。
原癌基因突变成癌基因,称为原癌基因的激活。
◆肿瘤抑制基因(tumor suppressor gene),抑癌基因(tumor suppressor gene) 两个拷贝,只有当两个拷贝都丢失了或两个拷贝都失活了才会使细胞失去增殖
的控制, 如:RB, P53
Harris(1968):癌细胞系与同组织正常细胞融合?杂交细胞无恶性表型,也不致
癌; 随着染色体丢失?则可能恢复致癌
肿瘤抑制基因与原癌基因突变对细胞的影响
特 点 基因属性 致癌方式
抑癌基因 原癌基因 细胞分化必需 细胞增殖必需 表达基因缺失或失活 基因突变激活、异常表达 loss of function gain of function